










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.48 n.2-3 San José Jun. 2000
Situación nomenclatural de Prionodactylus ampuedai
y Prionodactylus phelpsorum
Abraham Mijares-Urrutia 1
Recibido 13-V-1999 Corregido 21-I-2000 Aceptado 4-II-2000.
Abstract
New records of the poorly known microteiid lizard Prionodactylus ampuedai, morphological variation, and new data on natural history are reported. We report the loss of the holotypes of P. ampuedai and P. phelpsorum, and designate a neotype for the former. Euspondylus goeleti is resurrected from the synonymy with E. phelpsorum and this latter is transferred to the genus Prionodactylus.
Key words
Euspondylus phelpsorum, Prionodactylus ampuedai, Teiidae, Taxonomy, Squamata, Venezuela.
El género Prionodactylus está constituido por seis especies (Ávila-Pires 1995) pero, con las recientes adiciones de Gorzula & Señaris (1998), esta cifra se eleva a ocho. Tres especies se han registrado en Venezuela, P. ampuedae único representante del género en los Andes del extremo sudoeste del país y zonas adyacentes de los Andes de Colombia (La Marca & García-Pérez 1990), P. nigroventris y P. phelpsorum de las cimas de varios tepuyes de la guayana venezolana (Gorzula & Señaris 1998). La sistemática del género es problemática debido, parcialmente, a que Prionodactylus está definido con base en caracteres presentes en otros microtéidos. Según Ávila-Pires (1995) Pantodactylus y Prionodactylus no se diferencian claramente en su morfología externa, por lo que diagnostica el segundo género por tener cuerpo entre cilíndrico y ligeramente deprimido, redondeado en corte transversal; extremidades bien desarrolladas, pentadáctilos, todos los dígitos con uñas; escamas nasales separadas por una o dos frontonasales; párpado inferior con un disco semitransparente, escamas prefrontales, frontoparietales y occipitales presentes; interparietal más larga que las parietales; región gular con dos o más pares de filas de escamas agrandadas (éstas son ocasionalmente irregulares); dorsales hexagonales, alargadas, quilladas, dispuestas solamente en filas transversales; escamas laterales notablemente más pequeñas que las dorsales y ventrales lisas. Sin embargo, éstas características dificilmente implican que exista afinidad entre las ocho especies asignadas al género.
La taxonomía de Prionodactylus ampuedai y P. phelpsorum, permanece confusa e incierta. Ambas especies, desde que fueron descritas con base en un sólo ejemplar (Lancini, 1968), se han mantenido virtualmente ignoradas y la escasa información disponible en la actualidad es mayormente especulativa y/o anecdótica (Lancini 1968, Uzzell 1973, La Marca & García-Pérez 1990, Myers & Donnelly 1996, Gorzula & Señaris 1998). Prionodactylus ampuedai fue inicialmente descrito en el género Euspondylus, Uzzell (1973) sugirió su tranferencia al género Prionodactylus, agregando que esta pudiera ser conespecífica de P. vertebralis; más recientemente, La Marca & García-Pérez (1990) argumentan en favor de la validez del nivel de especíe para P. ampuedai proponiendo, además, la enmienda justificada del nombre de P. ampuedae a P. ampuedai. Por otra parte, Myers & Donnely (1996) proponen la enmienda justificada del nombre de E. phelpsi a E. phelpsorum y describen E. goeleti del Cerro Yaví, considerando ambas especies estrechamente relacionadas (Myers & Donnelly 1996: 30); Gorzula & Señaris (1998), proponen transferir sin explicaciones E. phelpsorum al género Prionodactylus y consideran a E. goeleti como un sinónimo reciente de E. phelpsorum.
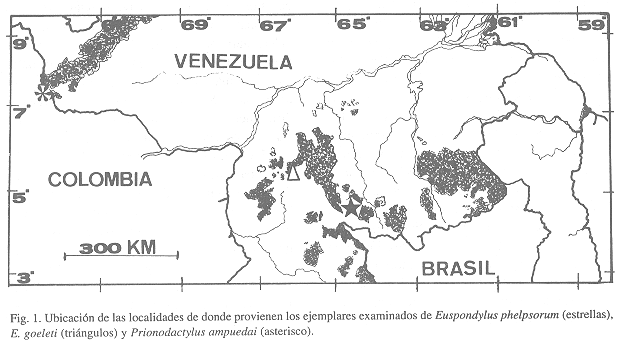
Para este estudio se localizaron 25 ejemplares adicionales de Prionodactylus ampuedai y se examinaron el holotipo y dos paratipos de E. goeleti más un ejemplar adicional del Cerro Yaví. El estudio de estos individuos permitirá hacer una primera aproximación de la variabilidad morfológica además de agregar nuevos datos de su historia natural (en el caso de P. ampuedai) y corroborar algunos de los cambios taxonómicos propuestos recientemente (en el caso de P. phelpsorum). Por otra parte, hacemos una revisión de la situación nomenclatural de P. ampuedai y E. phelpsorum.
Materiales y Métodos
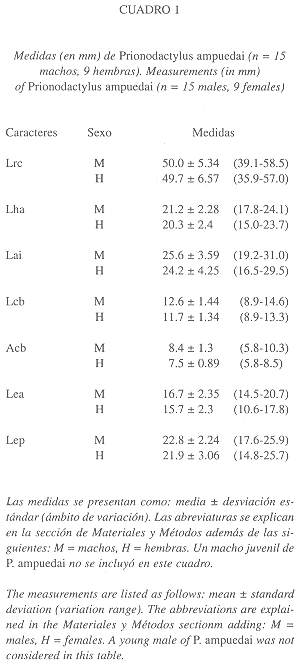
Seguimos la terminología y formato de descripción de Harris (1994). Las medidas se tomaron con un vernier (apreciación de ± 0.1 mm) bajo una lupa de disección, las medidas son las siguientes: longitud rostro-cloacal (desde la punta del hocico hasta el labio anterior de la cloaca, LRC), longitud hocico-axila (desde la punta del hocico hasta la cara posterior de la inserción de la extremidad anterior, LHA), longitud axila-ingle (entre la axila y la ingle de las extremidades del mismo lado, LAI), longitud cefálica (desde la punta del hocico hasta el margen anterior del oido, LHL), ancho cefálico (a nivel de la parte más ancha de la cabeza, ACB), longitud extremidad anterior (desde la axila hasta el extremo distal del dedo más largo, sin incluir la uña, LEA), longitud de la extremidad posterior (desde la ingle hasta el extremo distal del dedo más largo, sin incluir la uña, LEP). Para evaluar las diferencias en escamación, tamaño y proporciones entre ambos sexos empleamos un análisis de t-Student; por razones estadísticas, los caracteres de escamación con conteo bilateral se compararon como cifra total (se sumaron los conteos de izquierda y derecha). En el Cuadro 1 se resumen los datos merísticos de los ejemplares examinados de Euspondylus phelpsorum y Prionodactylus ampuedai. El sexo se determinó por disección. Todos los ejemplares examinados por nosotros fueron recolectados por Abdem R. Lancini y Ramón Ampueda en marzo de 1975 y por Julio Sánchez entre julio y agosto de 1975, sumando 25 ejemplares provienentes de dicha localidad (Fig. 1). Los ejemplares están depositados en las siguientes colecciones zoológicas (los datos de catálogo se detallan el el Apéndice): Estación Biológica de Rancho Grande (EBRG), Ministerio del Ambiente y de Los Recursos Naturales Renovables, Maracay; Fundación Museo de Ciencias Naturales de Caracas, sección de Herpetología (MCNC-HERP), Caracas.
Resultados
Lancini, 1968
Esta especie fue descrita con base en un único ejemplar proveniente del Cerro Jaua (Fig.1), catalogado en el MCNC. Myers & Donnelly (1996) modifican el nombre de Euspondylus phelpsi a E. phelpsorum. Sin embargo, no fue posible localizar el holotipo de Euspondylus phelpsorum (número MCNC 5269) y no existe ningún documento que señale su ubicación actual y, además, en el catálogo del MCNC no están registrados ejemplares adicionales de E. phelpsorum (Luis F. Navarrete, conservador de la sección de herpetología del MCNC, comunicación personal, 1998. El Dr. Abden R. Lancini, anterior conservador y director del MCNC, confirmó esta información).
Uzzell (1973) cuestionó la asignación genérica de E. phelpsorum por considerar inusual en Euspondylus la presencia de una línea blanca que se extiende entre las supralabiales posteriores y la inserción de la extremidad anterior, aunque el autor reconoce que este tipo de línea también aparece en E. simonsi y E. acutirostris. Nuestras observaciones de E. acutirostris no revela tal línea en las escamas supralabiales. Gorsula & Señaris (1998) emplean la combinación Prionodactylus phelpsorum sin explicaciones que justifiquen el cambio.
Recientemente, Gorzula & Señaris (1998) plantean que E. goeleti es un sinónimo reciente de E. phelpsorum. Ellos cuestionaron las características diferenciales dadas por Myers & Donnelly (1996) para E. goeleti, una línea mediodordal ensanchada posteriormente (contra unifórmemente estrecha a lo largo del tronco), coloración ventral diferente y las escamas dorsales notablemente quilladas (contra débilmente quilladas en E. phelpsorum). Del examen del holotipo de E. goeleti (EBRG 3112), dos paratopotipos (EBRG 3113-14) y un ejemplar adicional (EBRG 3035), se desprenden las siguientes observaciones: (1) La línea pálida medio dorsal es de 1-2 escamas de ancho en la nuca, 2½ -3 escamas a nivel de la inserción de las extremidades anteriores, ensanchándose progresivamente hasta alcanzar 4-6 escamas de ancho a nivel de la inserción de las extremidades posteriores; se destaca el ejemplar EBRG 3114 (un juvenil) en que la línea mediodorsal es algo difusa entre las escamas occipitales y la inserción de las extremidades anteriores y, a continuación, progresivamente se difumina hasta desaparececer a nivel de la mitad longitudinal del cuerpo. (2) La coloración de las escamas mentales y gulares es entre blanco cremoso con el centro gris plomiso hasta gris azulado con los bordes irregularmente blancos; las escamas ventrales entre crema con manchas oscuras algo difusas hasta gris azulado uniforme con el borde posterior de cada escama claramente más pálido. (3) Las quillas de las escamas dorsales son entre débil a claramente demarcadas, en ningún caso notablemente quilladas. Éstas observaciones sugieren que la sinonímia propuesta por Gorzula & Señaris (1996) estaría bien fundamentada. Sin embargo, debido a la pérdida del holotipo de E. phelpsorum (y la inexistencia de material adicional), no es posible resolver la identidad dudosa de E. goeleti en la descripción original (Myers & Donnelly 1996), consideramos que se debe retomar el nombre de E. goeleti de la sinonímia con E. phelpsorum. Para justificar el uso del nombre específico phelpsorum y facilitar la comparación con especies y poblaciones similares, queda pendiente la designación de un neotipo.
Por otra parte, E. goeleti y E. phelsorum presentan una línea blanca supralabial, escamas dorsales débilmente quilladas y hexagonales, tres escamas supraoculares, y un par de filas longitudinales de gulares agrandadas (al menos en E. goeleti), entre otras caracteres, lo que asocia claramente ambas especies con el género Prionodactylus (según Uzzell 1973, Ávila-Pires 1995). Sin embargo, dado que Prionodactylus no puede ser definido por caracteres derivados únicos, E. phelpsorum y E. goeleti son asignados tentativamente al género Prionodactylus como una cuestión de conveniencia y proponemos las nuevas combinaciones Prionodactylus goeleti (Myers & Donnelly 1996) y Prionodactylus phelpsorum (Lancini 1968).
Designación del Neotipo de
Prionodactylus ampuedai (Lancini 1968)
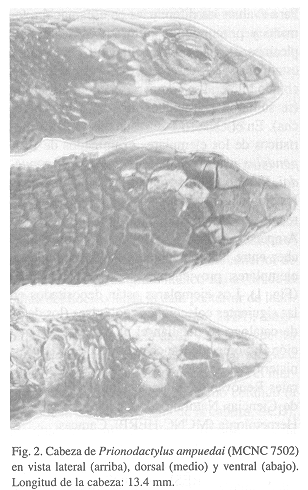
Figs. 2
Esta especie tiene los mismos problemas nomenclaturales que Euspondylus phelpsorum. El holotipo y cuatro paratipos (MCNC 5223, 5263, 5265, 5396 y 5398, respectivamente) se consideran actualmente perdidos (Luis F. Navarrete, conservador de la sección de herpetología del MCNC, comunicación personal, 1998. El Dr. Abden R. Lancini, anterior conservador y director del MCNC, confirmó esta información) y, para justificar el uso del nombre específico ampuedai y facilitar la comparación con especies y poblaciones similares, procedemos a designar un neotipo, según los lineamisntos del artículo 75 del Código de Nomenclatura Zoológica (1985). El neotipo de Euspondylus ampuedaeLancini 1968, al igual que el holotipo perdido, proviene de la localidad tipo de la especie y es una hembra adulta. Describimos este ejemplar y la variación vista en los restantes ejemplares (ver Apéndice) se incluye entre paréntesis.
Prionodactylus ampuedai (Lancini, 1968)
Euspondylus ampuedaeLancini, 1968: 4. Holotipo (MCNC 5223) y paratipos (MCNC 5265, 5263, 5396 y 5398) perdidos. Localidad tipo: ...en los alrededores de Villa Páez (Edo. Táchira, Venezuela) a una altura superior a los 2.000 metros sobre el nivel del mar.
Prionodactylus vertebralisUzzell, 1973: 7.
Prionodactylus ampuedaiLa Marca & García-Pérez, 1989: 112.
Neotipo: MCNC 7544, hembra adulta, proveniente de Villa Páez, Municipio Junín, estado Táchira, Venezuela (aproximadamente 7°29 N, 72°26 W), ± 2000 m, por Julio Sánchez, el 29 de julio de 1995.
Descripción: Escamas prefrontales en amplio contacto por la sutura media; escamas supraoculares 3 (rara vez 2); supraciliares 3 (rara vez 4); serie de supraciliares separan completamente las escamas supraoculares de las escamas ciliares; escama interparietal con los márgenes laterales casi paralelos; todas las escamas dorsales de la cabeza (excepto las temporales) lisas o ligeramente rugosas (sin crestas ni protuberancias); escamas temporales ligeramente quilladas; número de temporales 14 (izquierda, 14-21), 18 (derecha, 14-21); escamas occipitales más pequeñas que las temporales, pentagonales, paravertebrales; segunda fila postparietal ausente; escama nasal entera, estrecha anteriormente, casi pentagonal; narinas redondeadas, situada en el centro de la nasal, abertura dirigida latero-posteriormente, canal descendiente; escama loreal separada de las supralabiales por una escama freno-orbital anterior; concavidad ciliar apenas evidente; palpebrales superiores 3; palpebrales inferiores 3 (derecha, 1-3), 2 (izquierda, 1-3); palpebrales inferiores ligeramente manchadas, translúcidas; tres filas de gránulos debajo de las papebrales inferiores (1-5 filas); 5 (3- 6 por lado) escamas suboculares en contacto con las supralabiales; una fila de gránulos sobre el borde anterior del oido; gránulos preauriculares 7 (izquierda, entre 5-8), 6 (derecha, entre 5-7; gránulos preauriculares anchos; escamas supralabiales 7 (rara vez 6), sexta (o cuarta, a nivel de la inflexión del labio) tan ancha como alta; escamas infralabiales agrandadas 5 (4-6); escamas geneiales agrandadas una anterior seguida por 4 pares; par anterior de geneiales totalmente unidas por el medio, los restantes pares de geneiales separadas; escamas sublabiales bordean la cuarta infralabial y cuarta geneial; pregulares no agrandadas, dispuestas oblícuamente; pliegue gular evidente; fila transversal de escamas gulares 7 (6-8, incluyendo las escamas del collar); serie paramediana de escamas gulares presente, en fila doble de escamas agrandadas, cuadradas; escamas del collar 5 (4-10, normalmente entre 5-7), centrales más estrechas; escamas laterales del cuello ¼ (o hasta 1/5) del tamaño de las escamas de la nuca; pliegue del collar débil pero obvio, no contiene gránulos; pliegue gutural débil pero obvio.
Escamas alrededor del medio del cuerpo (a nivel de la fila transversal media de escamas ventrales) 27 (26-31); filas longitudinales de escamas dorsales 13 (11-16); filas de escamas laterales 4 por lado (3-6); filas longitudinales de escamas ventrales 6 (3-6); escamas laterales del cuerpo notablemente más pequeñas que las escamas dorsales y ventrales; filas transversales de escamas dorsales entre la escama occipital y el margen posterior de la inserción de las extremidades posteriores 36 (32-37); filas transversales de escamas ventrales entre el collar y el labio anterior de la cloaca 19 (18-22); escamas dorsales 1.7 (1.6-2.0) veces más largas que anchas; escamas dorsales con una quilla central, bien definida (no pronunciada), estrecha; escamas dorsales esencialmente hexagonales o con una ligeras convexidad redondeada en el margen posterior, subimbricadas o imbricadas, filas tranversales alternadas; escamas medioventrales cuadradas o rectangulares (cuando se fusionan), más estrechas hacia los lados, subimbricadas (rara vez yuxtapuestas), lisas; margen posterior recto en las escamas ventrales; escamas ventrales con las esquinas curvadas; escamas precloacales 4 (4 en todas las hembras, 2 en todos los machos); escamas redondeadas sobre el labio posterior de la cloaca; margen posterior de las escamas supracaudales truncado; escamas subcaudales con margen posterior recto.
Escamas dorsales de las extremidades rugosas; escamas supradigitales del IV dedo manual 8 (izquierda, 8-10), 9 (derecha, 7-10); escamas infradigitales del IV dedo manual 13 (por lado, 12-16); algunas escamas infradigitales proximales tuberosas, sencillas, la mayoría divididas por el medio; escamas bajo el pólex de la mano 5 (izquierda), 4 (derecha) (4-7 por lado); escamas en palmas de mano y pies tuberosas, no cornificadas ni agrandadas; escamas supradigitales del IV dedo pedal 11 (izquierda, 9-13), 10 (derecha, 9-12); escamas infradigitales bajo el IV dedo pedal 18 (izquierda, 18-20), 19 (derecha, 17-22); algunas escamas infradigitales proximales sencillas y tuberosas, la mayoría divididas por el medio; uña de tamaño mediano (aproximadamente 1/2 del tamaño de la escama adyacente o casi tan grande como la escama adyacente); porción distal de los dígitos ligeramente comprimidos a los lados; escamas poríferas enteras; poros preanales ausentes; poros femorales 6 (por lado, poros femorales en hembras 4-8 [derecha], 3-9 [izquierda]; machos 9-11); extremidades del mismo lado se superponen +6 escamas cuando son estiradas hacia el centro del cuerpo (entre +3 a +10 escamas); dígitos en orden decreciente de longitud 4 > 3 > 2 = 5 > 1.
Superficie dorsal de la lengua cubierta por papilas en forma de escamas, dispuestas en filas oblícuas; pliegus infralinguales estrechos, oblícuos; dientes bicúspides, unos pocos dientes frontales cónicos, 20 (16-23, n = 13) a lo largo de de la mandíbula superior, 17 (15-19, n=13) a lo largo de la mandíbula inferior; paladar con hendidura media.
Coloración en preservativo: Dorso castaño oscuro, excepto en 1/3 de los ejemplares examinados (MCNC 7494, 7545-49, 7504-05) presentan una línea mediodorsal, ancha, ligeramente más pálida que el dorso (entre casi indistinta y bien visible), desde la cabeza hasta la cola. Vientre igual que el dorso excepto el borde posterior de las escamas ventrales el cual es crema sucio, éste último progresivamente más extenso desde las ventrolaterales hacia las escamas medioventrales. Una línea blanca desde la primera supralabial hasta el borde anterior del oido, pero frecuentemente (en alguno de los lados de no menos de 17 ejemplares) esta línea alcanza por delante (o hasta el nivel) de los hombros; en el lado izquierdo de la cabeza del ejemplar MCNC 7507, esta línea alcanza por detrás de los hombros. Superficie ventral de las extremidades castaño pálido; cuatro ejemplares (MCNC 7410, 7494-95, 7507) con el mentón y región pregular castaño pálido; vientre grís castaño en MCNC 7545.
Comparaciones: Dentro del género Prionodactylus, P. ampuedai se distingue por poseer una línea blanca supralabial excepto de P. dicrus, P. manicatus, P. nigroventris, P. phelpsorum y P. vertebralis (según Uzzell 1973). Tres especies, P. ampuedai, P. dicrus y P. vertebralis son únicas en tener la escama loreal separada de las supralabiales por la escama nasal y la freno-orbital y una línea pálida mediodorsal. El número de poros femorales se ha registrado como sexualmente dimórficos en tres especies P. dicrus y P. argulus (Uzzell 1973) y P. ampuedai. Prionodactylus ampuedai difiere de P. vertebralis por tener 27-31 escamas alrededor del medio del cuerpo (31-45 en P. vertebralis) y presentar dimorfismo sexual en el número de poros femorales (el número de poros femorales no es dimórfico en P. vertebralis); difiere de P. dicrus por presentar una única línea pálida mediodorsal (dos líneas entre el hocico y la porción anterior del cuerpo en P. dicrus) y por tener 3-9 poros femorales en las hembras y 9-11 en los machos (7-11 en las hembras y 20-25 en los machos de P. dicrus).
Distribución e historia natural: Actualmente, Prionodactylus ampuedai ha sido registrado sólo en la localidad tipo en el extremo sudoccidental de los Andes de Venezuela y de una localidad adyacente en los Andes colombianos (La Marca & García-Pérez 1990). La localidad venezolana se sitúa dentro de la asociación vegetal descrita como Bosques submontanos y montanos ombrófilos según Huber & Alarcón (1988). No se dispone de información sobre el microhábitat ni condiciones reproductivas.
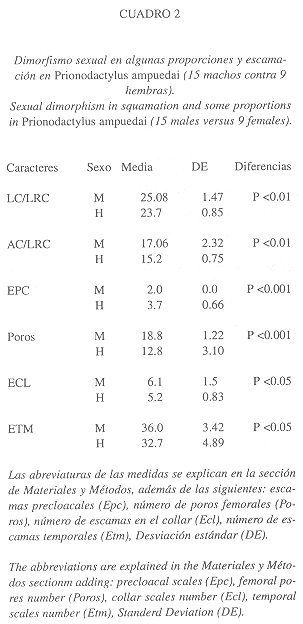
Dimorfismo sexual: Cuando menos seis caracteres han mostrado ser sexualmente dimórficos en Prionodactylus ampuedai, dos de escamación y tres de tamaño o proporciones (ver Cuadro 2). El número de escamas precloacales y número de poros femorales permite distinguir a primera vista los machos de las hembras; los machos poseen dos escamas precloacales y un número elevado de poros femorales (9-11), contra cuatro escamas precloacales y un menor número de poros femorales (3-9) en las hembras.
Ejemplares examinados: Todos los ejemplares en esta lista provienen de localidades dentro de Venezuela. Prionodactylus ampuedai: Táchira: MCNC 7410, 7427, 7429, 7494-95, 7499, 7544, 7504, 7547 (hembras), 7428, 7493, 7496-98, 7500-03, 7505-07, 7545-46, 7548-49 (machos), Villa Páez, Municipio Junín. Euspondylus goeleti: Amazonas: EBRG 3112 (holotipo), hembra, EBRG 3113-14 (paratipos), juveniles, Cima del Cerro Yaví, 2150 m.- EBRG 3035, macho, Cerro Yaví, 2100 m.
Agradecimientos
Gracias a Luis F. Navarrete (MCNC) por la valiosa información sobre los holotipos de P. ampuedai y E. phelpsorum. Celsi Señaris facilitó un mapa a partir del cual Nadhia Rujana preparó la Figura 2. Dos revisores anónimos hicieron valiosas sugerencias sobre el manuscrito. Este trabajo se presentó en la XIII Reunión de Comunicaciones Herpetológicas, Santa Fe, Argentina, Septiembre de 1998, con la subvención del CONICIT, expediente N° S2-98001514.
Referencias
Anónimo. 1985. International Code of Zoological Nomenclature. International Trust for Zoological Nomenclature, Londres. [ Links ]
Ávila-Pires, T. C. S. 1995. Lizards of the Brazilian amazonia. Zool. Verh. Leiden 299: 1-706. [ Links ]
Gorzula, S. & J. C. Señaris. 1998. Contribution to the herpetofauna of the venezuelan Guayana I. A data base. Scientia Guyanae 8: 1-270. [ Links ]
Harris, D. M. 1994. Review of the teiid lizard genus Ptychoglossus. Herp. Monogr. 8: 226-275. [ Links ]
Huber, O. & C. Alarcón. 1988. Mapa de Vegetación de Venezuela. IUCN-Ministerio del Ambiente y los Recursos Naturales Renovables, Armitano, Caracas. [ Links ]
La Marca, E. & J. E. García-Pérez. 1990. Prionodactylus ampuedai (Sauria: Teiidae): a new lizard record for Colombia, with taxonomic comments on the species. Bull. Maryland Herp. Soc. 26: 111-115. [ Links ]
Lancini, A. R. 1968. El género Euspondylus (Sauria: Teiidae) en Venezuela. Publ. Ocas. Mus. Cienc. Nat. Caracas, Zool. 12: 1-6. [ Links ]
Myers, C. W. & M. A. Donnelly. 1996. A new herpetofauna from Cerro Yaví, Venezuela: first results of the Robert G. Goelet American Museum-Terramar Expedition to the northwestern tepuis. Amer. Mus. Novitates 3172: 1-56. [ Links ]
Uzzell, T. 1973. A revision of lizards of the genus Prionodactylus, with a new genus for P. leucostictus and notes on the genus Euspondylus (Sauria, Teiidae). Postilla 159: 1-67. [ Links ]
1 Colección Herpetológica Regional, Centro de Investigaciones en Ecología y Zonas Áridas (CIEZA),Universidad Francisco de Miranda, Apartado 7559, Santa Ana de Coro 4101-A, Venezuela. Fax: +58-68-524922. Correo-electrónico: amijares@funflc.org.ve

