











 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.48 no.2-3 San José Jun. 2000
and mating systems of L. surculus and L. byssiseda
Ronald H. Petersen 1
Received 9-XII-1998. Corrected 19-XI-1999. Accepted 30-XI-1999.
Abstract
The following names are proposed for species of Lentaria new to science: L. boletosporioides from New Zealand with stout basidiomata and boletoid basidiospores; L. caribbeana from Caribbean islands and circum-Caribbean landmasses, with large basidiomata and large basidiospores; L. glaucosiccescens from New Zealand, with basidiomata turning grey-green upon drying; and L. rionegrensis from Argentina, with densely branched, ruddy basidiomata. Lentaria javanica nom. nov. is proposed to substitute for Clavaria compressa, a later homonym. Basidiospore statistics indicate infraspecific differentiation within L. surculus and two morphological variants are described. Lentaria surculus and L. byssiseda exhibit tetrapolar mating systems, and the two species are sexually interincompatible
Key words
Biogeography, mating systems, new species, Lentaria.
Corner (1950) proposed the genus name Lentaria to accommodate two groups of taxa, both characterized by coralloid, pliant, often tough basidiomata on dead wood, white spores and clamped hyphae. The two groups were: 1) branched basidiomata often with basidiospores exhibiting high length-to-width ratios (i.e. E or Em values), including the type species, L. surculus; and 2) usually unbranched basidiomata of small size, usually lichenized with algae. Petersen (1967) segregated the latter group into the genus Multiclavula, leaving the type complex of Lentaria more homogeneous.
For the coralloid group, Corner (1970) introduced two subgenera: 1) subg. Lentaria (as Eu-lentaria), which by including L. surculus took in the non-phycophilous species with generally branched basidiomata; and 2) subg. Lentariopsis, with L. epichnoa as its type, intended to gather several anomalous taxa such as L. afflata (Lagger.) Corner and L. delicata (Fr.) Corner. In a discussion of the relationships of Lentaria (Corner 1970: 228), Corner pointed to Ramaria subg. Lentoramaria as a likely ally, also concluded by Petersen (1971a).
Petersen (1972) examined numerous specimens of Clavariadelphus ligulus (Fr.) Donk and C. sachalinensis (Imai) Corner, and Petersen (1974c) described C. lignicola Pet., which combined the long spores of Clavariadelphus subg. Ligulus with the lignicolous habit and branched basidiomata of Lentaria. Thus I now consider Lentaria, as typified by L. surculus, to be most closely allied to Clavariadelphus, which in turn is pivotal as the link between Gomphus-like taxa (tan, roughened spores; cantharelloid habit) and clavarioid taxa (i.e. Ramariopsis, particularly subg. Laevispora).
Inventorying in Costa Rica in 1995 and 1998 yielded several variant specimens, supplementing examination of fresh collections and herbarium material of Lentaria taxa over the years. Some new taxa have been identified and they are proposed below, together with a discussion of infraspecific variation in L. surculus.
Materials and Methods
Colors enclosed in quotation marks are from Ridgway (1912), while those cited alphanumerically are from Kornerup and Wanscher (1967). Single-basidiospore isolates were established following methods described by Petersen and Gordon (1991). E = spore length divided by spore width; Em = mean spore length divided by mean spore width; Lm = mean spore length.
Results
R.H. Petersen, sp. nov.
Basidiocarpia terrestris, ramosa, ad 6 cm alta, gregaria ad caespitosa; stipite et ramosa cinnamomea; apices elongato, aceroso, albidior. Mycelium basalis albis, tomentosis, cum rhizomorphae albae.
Hyphae basalis fibulatis, 2.5-6 µm lato. Hyphae ramosae 2-3.5 µm lato, fibulatae, tenuitunicatae. Basidia 60-80 µm longa, clavata, fibulata. Basidiosporae 15-24 X 4.5-6 µm, sigmoideae, tentuitunicatae.
Holotypus: Novo Zealandeae, Marlborough, Totara, Pelorus Scenic Preserve, 19.V.82, TENN 43133.
Basidiomata up to 6 cm high, up to 2 cm broad, branched in 1-3 ranks, gregarious to densely cespitose, arising from extensive white to ivory-colored ("cartridge buff;" 30A2) resupinate patches and rhizomorphs on well-rotted wood, off-white when young, slowly becoming fleshy tan, cinnamon to dull avellaneous ("avellaneous," "tilleul-buff," "cinnamon," "clay-color," "cinnamon-buff," "cream-buff;" 7B2-3, 6B4-5, 5C6). Stipe discrete, flattened to lobed in cross-section, tan. Branches flattened in cross-section; axils acute; internodes long below, diminishing gradually. Apices somewhat paler than branches ("light pinkish cinnamon;" 7A2), prolonged, awl-shaped; flesh drying light in weight and friable. Taste faintly bitter; odor negligible when fresh, mild of fenugreek when dry.
On rotting wood or bark chips, including Nothofagus.
Hyphae of resupinate patch 2.5-6 µm diam, thin- to thick-walled (wall up to 0.6 µm thick), conspicuously clamped, smooth, of two types: 1) contents staining weakly in cotton blue, composing the cortical areas of rhizomorphs and all of the resupinate patches; and 2) contents staining strongly in cotton blue, limited to medullary portions of rhizomorphs. Upper branch tramal hyphae narrow, 2.0-3.5 µm diam, thin- to thick-walled (wall less than 0.5 µm thick), often minutely distinctly banded or papillose under phase contrast microscopy, conspicuously clamped, tightly packed, parallel. Subhymenium extensive, undifferentiated; hyphae 1.5-3.0 µm diam, thin-walled, clamped. Basidia 60-85 X 8-11 µm, clavate, attenuate below, clamped, multiguttulate when mature; sterigmata 4, long, curved-divergent.
Spores 15-24 X 4.5-6 µm, boletoid, sway-back, to gently sigmoid, smooth, thin-walled, hyaline, non-reactive in IKI; contents multiguttulate when mature; hilar appendix gradual, not prominent.
Specimens examined: NEW ZEALAND, North Island, Urewera National Park, Aniwaniwa Falls Track, 26.V.81, coll. & det. RHP, TENN 42202; South Island, Marlborough, Totara, Pelorus Scenic Reserve, 19.V.82, coll. & det. RHP, TENN 43133 (HOLOTYPE); Same location, 19.V.82, coll. & det. RHP, TENN 43138; same location, Totara Trig Track, 21.V.82, coll. & det. RHP, TENN no. 43132; 5 km W of Reefton, vic. rte 7, 11.VI.81, coll. & det. RHP, TENN 42247.
Commentary: Petersen (1988) included two groups of specimens from New Zealand under the name Lentaria surculus. One group, cited above, represents material of L. boletosporioides, while some others are truly L. surculus (listed below). The two species, as represented by New Zealand material, can be separated as follows: L. boletosporioides produces stouter, erect basidiomata which rarely branch in more than 2 ranks; except for branch apices, all basidiome parts are usually strap-shaped; basidiospores are boletoid, sway-back to gently sigmoid; upper branch tramal hyphae are noticeably banded or papillose under phase microscopy. Lentaria surculus produces more delicate, often decumbent basidiomata, usually branched in three or more ranks; all basidiome parts are typically terete; basidiospores are straight or nearly so; upper branch tramal hyphae are smooth under phase microscopy. It was L. surculus which was figured by Petersen (1988: fiche 1, pl. 53), but text fig. 67 depicted spores of L. boletosporioides.
New Zealand specimens of L. surculus examined are as follows: NEW ZEALAND, North Island, Waipoua Kauri Reserve, Big Kauri Tree Track, 23.VI.81, coll. & det. RHP, TENN no. 42263; Puketi State Forest, 2.VI.82, coll. G. Samuels, det. RHP, TENN 43445; Waipoua Kauri Reserve, Yakas Tree Track, 12.V.83, coll. G. Samuels, det. RHP, TENN 43815.
Lentaria caribbeana
R.H. Petersen, sp. nov.
Basidiocarpia terrestris, ramosa, robusta, ad 12 X 6 cm. Tomentum basalis alba, extensiva; hyphae 3-5 µm lato, crassitunicatae, fibulatae. Stipite ad 2 X 1.5 cm, pallide brunnea, ad 4-7-plo e basi sursum ramificans. Ramosa brunnea ad cinnamomea. Apices acerosa, albidior. Basidia 23-28 X 6-7 µm, fibulata, clavata. Basidiosporae 7.6-9.1 X 2.9-3.6 µm, ellipsoidea ad cylindracea, tentuitunicatae.
Holotypus: Costa Rica, Prov. Puntarenas, Canton Coto Brus, Hacienda La Amistad, 18.VI.95, TENN 53842.
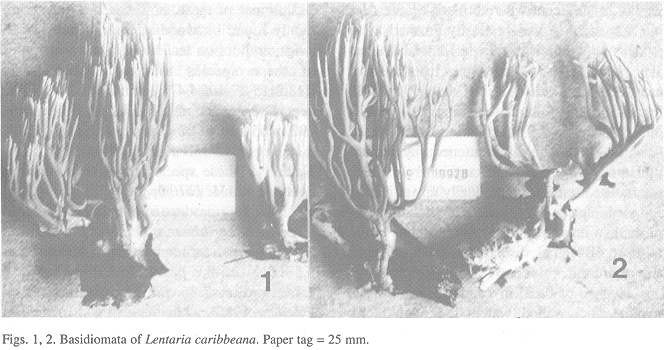
Basidiomata (Figs. 1, 2) up to 12 X 6 cm, repeatedly branched, obovate in outline, arbuscular. Basal mat usually extensive, comprising two tissues: 1) superficial, tough-tomentose on subtratum surface, white, usually discretely bounded; and 2) amorphous within substratum, but with slender innate rhizomorphs, often extensively binding litter, white to cream-colored, unchanging when handled. Stipe usually discrete, single (not cespitose), up to 2 X 1.5 cm, irregularly rounded, orange-ochraceous brown ("ochraceous tawny" to "buckthorn brown;" 6C6, 5D6), often covered by a felty mycelial layer; stipe flesh off-white, felty to punky, somewhat watery brunnescent where bruised or cut, drying very light in weight, easily crushed, with evidence of dimidiate growth. Major branches 3-6, more or less erect, somewhat flattened, cream-colored below, beige or tan upward; branching dichotomous to sympodial throughout; branches in 4-7 ranks, more or less erect, substrict, beige or tan, flattened; hymenium unilateral, sterile areas decurrent from axils; internodes diminishing gradually upward; axils rounded; apices awl-shaped to dichotomous, paler than branches ("chamois" to bone white at tips; 4B4). No discernable color changes where bruised. Odor and taste negligible.
Hyphae of basal mycelium 3-5 µm diam, hyaline, thick-walled (wall up to 0.5 µm thick), conspicuously clamped, straight, parallel, not adherent; walls weakly cyanophilous; ampulliform swelling usually as inflated clamp connections, occasionally as abrupt, lateral, subbulbous hyphal protuberances, not unusually thick-walled, unornamented, usually elongate barrel-shaped; echinulate hyphae not observed. Tramal hyphae of upper branches 4-12 µm diam, hyaline, thick-walled (wall up to 1 µm thick), conspicuously clamped, more or less straight, parallel, not agglutinate or adherent; ampulliform clamps and gloeoplerous hyphae not observed. Subhymenium extensive, pseudoparenchymatous. Hymenium thickening; basidia 23-28 X 6-7 µm, clavate, clamped, yellowish and somewhat refringent under phase contrast; contents homogeneous or with minute granules or guttules, especially proximally; sterigmata 4, slender, more or less straight. Leptocystidia 22-30 X 2.5-4 µm, hyphal, occasionally lobed or angled, exerted up to 7 µm from hymenium; contents homogeneous.
Spores 7.6-9.1 X 2.9-3.60 µm (E = 2.40-2.89; Em = 2.67; Lm = 8.52 µm), rhomboid to ellipsoid, usually with suprahilar inflation ("boletoid"), hyaline, thin-walled; contents minutely heterogeneous to 1-3-guttulate; hilar appendix stout, blunt, not prominent.
Commentary: Only L. byssiseda (Pers.: Fr.) Corner (typified by Petersen, 1989) and its tropical analog, L. surculus (Berk.) Corner approach this taxon in basidiome size. Basidiomes of these, however, are most commonly found on woody debris, usually small twigs, and not on leaf litter. Moreover, spores of these species are appreciably larger [(13.2-)15-17.4 X 4.4-5.2 µm; E = 2.77-3.81 for neotype specimen of L. byssiseda, cf. Petersen, 1972; 15-24 X 4.3-6 µm for New Zealand material of L. surculus, cf. Petersen, 1988]. Neither of these species could be considered contaxic with L. caribbeana.
Two taxa produce spores quite similar to those of L. caribbeana. Lentaria patouillardii (Bres.) Corner spores have been measured as 6.5-8.5 X 2.5-3.2 µm (E = 2.56-3.14; Em = 2.74; Lm = 7.86 µm, for lectotype, cf. Petersen, 1985), while those of Clavaria micheneri B. &. C. were measured as 6.5-7.6 X 4.0-4.3 µm (E = 1.64-1.91; Em = 1.77; Lm = 7.2 µm, for lectotype, cf. Petersen, 1985). The latter, known to me from only a few specimens, forms basidiomes up to 6 cm high, superficially like those of the Puerto Rico taxon, but its spores are more like those of L. pinicola (Burt) Pet. in shape (for the holotype of the latter, 7.7-9.3 X 4.5-5.7 µm; for the taxon 7.5-10.9 X 4.2-5.9 µm, cf. Petersen, 1974b), being short-cylindrical to broadly ellipsoid. Thus, L. patouillardii bears spores most similar to those of the Puerto Rico specimen.
My experience indicates that L. patouillardii fruits in temperate (not subtropical or tropical) forests, from northern Italy (the only European location to my knowledge) through temperate North America. No basidiomata larger than 4 cm high have been seen, however, and these are always grayish beige (like dried material of Clavulina cristata). The taxon fruits on woody debris, especially small twigs. With these differences in basidiome morphology, habitat and distribution, I am obliged to keep L. patouillardii separate from L. caribbeana, in spite of spore similarities.
Other, less significant characters distinguishing L. caribbeana include basidiome color, which seems well within the range of the genus [most taxa tan, beige, gray-tan, with basidiomata of L. epichnoa (Fr.) Corner virtually totally white]. Presence of leptocystidial elements cannot be discounted, but they are inconspicuous, and probably represent basidioles.
Other taxa reviewed as possibly contaxic with L. caribbeana include Clavaria cervina B. & C. (lectotype, Fungi Cubensis Wrightiana, Nouvelle Sophia, 2.IX, Wright no. 235, K; synlectotype, FH, and presumably other distributions) and C. pallida B. & C. (lectotype, Fungi Cubensis Wrightiana, Cuba, 14.VIII, Wright no. 256, K; synlectotype, FH, and presumably other distributions). In my opinion, both are synonymous under L. surculus.
It might be noted that Clavaria compressa Berkeley [1842. Ann. Mag. Nat. Hist. 10: 383; = Lachnocladium compressum (Berk.) Léveillé. 1846. Ann. Sci. Nat., Ser. 3: 153] is a later homonym of C. compressa Schweinitz (1832. Trans. Amer. Phil. Soc. II 4: 182.), and therefore requires a new name. The epithet is based on a type specimen from Java, and I offer Lentaria javanica as a nomen novum.
Specimens examined (L. caribbeana): COSTA RICA, Prov. Punta Arenas, Canton Coto Brus, Hacienda La Amistad, 8o 54' 52" N, 82o 46' 50" W, 18.VI.95, coll. RHP, on hardwood debris, no. 7853 (TENN no. 53842; HOLOTYPE); same location, 19.VI.95, coll. RHP, no. 7858 (TENN no. 53834); same location, 5.VII.98, coll. RHP, no. 9927, 9928 (TENN 56553, 56554). UNITED STATES, Florida, Dade Co., Everglades National Park, Gumbo Limbo Trail, 7.VIII.72, coll. H.H. Burdsall, no. 6960 (TENN no. 38409); Puerto Rico, Luquillo Experimental Forest, El Verde Field Station, 10.VIII.88, coll. D.J. Lodge, s.n. (TENN); Puerto Rico, location unknown, 14.VI-22.VII.01, coll. L.M. Underwood & R.F. Griggs, no. 1002 (NY; TENN no. 32270).
Specimens examined (L. patouillardii): Canada. Manitoba, Winnipeg, 26.vii.28, Bisby no. 4221 (NCU); same location, 5.X.27, Bisby no. 4223 (NCU). Italy. Monte di Cappucino, X.1891, Bresadola, s.n. (lectotype, S; merotype, NCU); same location, XI.00, Bresadola, s.n. (NY). United States: California; Tahoe National Forest, 8.VI.86, coll. H. Saylor, no. 47015 (TENN). Idaho; Kootenai Co., Priest Lake, 25.IX.68, nos. 33730, 33740 (TENN); same location, 27.ix.68, nos. 33731, 34186, 34188, 34214 (TENN); same location, 3.X.68, no. 38710 (TENN); Boulder Creek, 4.VIII.66, coll. P. Miller, no. 40552 (TENN); Priest Lake, 19.IX.68, no. 33723 (TENN). Indiana; Brown County Park, no date, coll W.B. Cooke, no. 31539 (TENN). Iowa; Jefferson City, 7.VIII.53, coll. unknown, nos. 33363, 33376, 33650 (TENN). Michigan; Harbor Springs, 14.x.32, Mains no. 32-783 (NCU). Minnesota; Crow Wing Co., Pelican Twp., 9.IX.68, Weaver no. 1709, no. 31880 (TENN); same location, 16.VIII.68, Weaver no. 1678, no. 31883 (TENN). New York; Washington Co., Vaughn's Schoolhouse woods, 17.VII.16, Atkinson no. 28690 (CUP); same location, 16.VII.17, Atkinson no. 28328 (CUP); same location, 13.VIII.17, Atkinson no. 22534 (CUP); same location, 27.VII.17, Burnham no. 79 (NCU). Wisconsin; Sister Bay, 18.IX.47, coll. Fensholt, ex herb. Doty, s.n. (NY).
Lentaria glaucosiccescens
R H. Petersen, sp. nov.
Basidiocarpia terrestris, ad 2 X 1.5 cm, fasciculata, ramosa, delicata, ad aurantio-brunnea contusa, pallide caetuleo-viridae siccescens. Tomentum basalis absentia, vel rhizomorphae albae. Stipite absentia. Ramosa delicata, fusco-pallide ad pallide viride-brunnea. Basidia 55-60 X 9-10 µm, lato clavata, fibulatae. Basidiosporae 13-16.2 X 4.3-5.0 µm, sigmoideae ad cylindraceae, tenuitunicatae.
Holotypus: New Zealand, North Island, Auckland, Mill Bay, 6.VII.81, Horak num. 1059 (ZT; isotypus TENN 43612).
Description: Petersen, 1988: 77, fiche 1, pl. 54, text fig. 68. In addition, the following characters are noted. Branches colored 6C4 (near "avellaneous") to 6D4 ("buffy brown"). Taste and odor negligible.
Specimens examined: New Zealand, North Island, Auckland, Mill Bay, 6.VII.81, coll. E. Horak, Horak no. 1059 (holotype ZT; isotype, TENN 43612); South Island, Buller Dist., Cobb Valley, Asbestos Hut Track, 21.V.94, coll. A.S. Methven, det. RHP, on slender roots of tree fern rhizome, TENN no. 53676.
Commentary: Collection of a second specimen of this small, delicate fungus confirmed data from the first collection, which was reported by Petersen (1988). The pallid greenish blue shades upon drying are unique, and the fungus is easily overlooked due to its small size and depauperate appearance. Also unique are the delicate snow white rhizomorphs which remain so on drying, usually a characteristic of spiny-spored Ramaria taxa. In Lentaria, subiculum and rhizomorphs usually become off-white to ivory-colored upon drying.
Lentaria rionegrensis
R.H. Petersen, sp. nov.
Basidiocarpia terrestris, ad 6 X 5 cm, robusta, ad quadruplo e basi sursum ramificans, cum tomentum basalis. Stipite robusto, compresso, fusco-violaceo. Ramosa compressa, robusta, pallide violacea. Apices comressa, cristata, pallidior ad albidior. Basidia 50-57 X 8-11 µm, clavata fibulata. Basidiosporae 15.2-17.6 X 4.8-5.6 µm, sigmoideae, tenuitunicatae.
Holotypus: Argentina, Prov. Neuquen, Parque Nacional de Lannin, 19.V.96, TENN 55058.
Basidiomata up to 6 cm high, up to 5 cm broad, branched in -4 ranks, arising from buried wood. Rhizomorphs and subicular mat white, tough, extensive (binding litter up to 8 cm from basidiome base), amorphous but with distinct innate rhizomorphs up to 2 mm thick. Stipes usually numerous, congested, arising from extensive basal mat, usually flattened, violaceous gray ("avellaneous;" 7B3). Lower and middle branched flattened, somewhat paler than stipe ("vinaceous buff;" 9B2). Apices flattened, cristate to irregular, pallid pinkish gray ("tilleul buff;" 7B2). Flesh off-white, very tough. Odor faintly fresh; taste none, consistency leathery.
Hyphae of rhizomorphs and basal mat 3-4 µm diam, thick-walled (wall up to 0.4 um thick), with common conspicuous clamp connections, tightly packed; crystals abundant, more or less spherical, spiculate, deposited between hyphae, easily disarticulated into minute cuneiform to triangular shards. Branch tramal hyphae 2-3.5 µm diam, hyaline, thick-walled (wall up to 0.4 µm thick), conspicuously and commonly clamped, occasionally locally swollen up to 8 µm at clamp connection, tightly packed, more or less parallel, adherent. Subhymneium rudimentary, undifferentiated; hyphae 2-3 µm diam, clamped. Hymenium thickened; basidia 50-57 X 8-11 µm, clavate, clamped, thin- to thick-walled (wall up to 0.4 µm thick), multiguttulate when young, less so by maturity; sterigmata four, up to 8 µm long, subcornute. Cystidia none.
Basidiospores (11.2-)15.2-17.6(-18.4) X 4.8-5.6 µm [E = (2.00-)2.85-3.50; Em 3.11; Lm = 15.91 µm], sigmoid to sway-back, smooth, hyaline, multiguttulate; wall thin; hilar appendix gradual, eccentric.
Commentary: Corner (1950, 1970) included only two species of Lentaria with spores as large as those of L. rionegrensis, L. byssiseda and L. surculus. Over the years, these two species have been separated only with difficulty, and the major portion of this paper is intended to shed light on this problem. Corner (1957) examined two of Singer's specimens, treating them under L. surculus. Corner (1957) stated: "Both Singer's collections have spores up to 17 X 5 µm, and the branch-tips tended to be cristate, which is unusual." The latter character, basidiome color, and spore shape are the three leading separating characters for L. rionegrensis.
In my experience, basidiomata of L. byssiseda and L. surculus exhibit terete stipes and branches, and awl-shaped apices. Lentaria byssiseda seems to fruit preferentially on standing trees, while L. surculus fruits on small dead branches on or near the ground. In neither case is the basal mat as ill-defined or as extensive as in L. rionegrensis. Moreover, although there is often a suggestion of sigmoid profile of basidiospores in L. byssiseda and L. surculus, spores of L. rionegrensis are consistently sigmoid, and often stouter distally than proximally.
This is surely what Singer (1969) included as L. surculus in the South American mycota, following Corner's (1950) broad treatment under that name. Two of the three Singer specimens were collected quite close to those listed below, while the third came from high elevation on the Chilean side of the Andes but at the same approximate longitude.
Specimens examined: ARGENTINA, Prov. Nequen, Parque Nacional de Lannin, Pto. Hua Hum, 19.V.96, coll. RHP, on buried Nothofagus wood beneath Chusquea, no. 8386 (TENN no.55058; HOLOTYPE); PARATYPES: ARGENTINA, Prov. Rio Negro, Parque Nacional de Lago Puelo, north shore of Lago Puelo, 10.V.96, coll. RHP, on Nothofagus debris, no. 8341 (TENN no. 55015); ARGENTINA, Prov. Chubut, Parque Nacional de los Alerces, south shore of Lago Rivadavia, 8.V.96, coll. RHP, on half-buried Nothofagus wood, no. 8319 (TENN no. 54995).
Lentaria surculus
(Pers.: Fr.) Corner
Corner (1970) used the name Lentaria soluta (Karsten) Pilát for the concept he (Corner 1950) previously had treated under L. byssiseda. Why the former name was taken up is unknown to me, for Clavaria byssiseda, sanctioned by Fries (1821: 476), and represented by a specimen in herb. Fries (UPS) typical of sensu Corner (Petersen, 1971) surely secures the oldest (and sanctioned) use of the epithet. Corner (1950), however, designated L. byssiseda nom. nov. (not comb. nov. as might be expected). Donk (stated by Corner as "in litt.") assumed that the Fries concept was that of Clavaria himantia, probably through Fries's use of that descriptor in his diagnosis of Clavaria byssiseda, but no other support for that interpretation seems to exist, and I prefer to resort to the sanctioned epithet (and transfer) Lentaria byssiseda (Fr.) Corner.
Petersen (1989) reviewed the nomenclature of Lentaria byssiseda (Pers.: Fr.) Corner, typified the name (Petersen, 1972, 1989) and described tissue culture mat characters (Petersen, 1974a), so that species can serve as a baseline for comparison with other putative taxa in the genus.
Conversely, Lentaria surculus, apparently quite similar to L. byssiseda, has not been adequately circumscribed in spite of Corner's (1950, 1970) classic contributions to taxonomic understanding of clavarioid fungi and Petersen's contribution (1972, 1989).
Lentaria surculus (Berk.) Corner. 1950. Ann. Bot. Mem. 1: 444.
Basionym: Clavaria surculus Berkeley. 1842. Hooker's J. Bot. 1: 154. [!]
- = Clavaria compressa Berkeley. 1842. Ann. Mag. Nat. Hist. 10: 383 [non Schweinitz. 1832] [!]
- = Lachnocladium compressum (Berk.) Leveille. 1846. Ann. Sci. Nat., III: 153.
- = Clavaria cervina Berkeley & Curtis. 1868. J. Linn. Soc. Bot. 10: 338. [!]
- = Lachnocladium cervinum (B. & C.) Patouillard. 1886. Journ. Bot. 3: 26.
- = Clavaria pallida Berkeley & Curtis. 168. J. Linn. Soc. Bot. 10: 338. [!]
- = Lachnocladium pallidum (B. & C.) Patouillard. 1886. Journ. Bot. 3: 26.
- = Lachnocladium tonkinense Patouillard. 1891. Journ. Bot. (Morot) 5: 314. [!]
Hyphae of rhizomorphs hyaline, thick-walled (wall up to 0.5 µm thick), conpicuously clamped, relatively straight, not agglutinated or adherent, tightly packed, involved in copious crystalline deposits; medullary hyphae smooth, with common ampulliform clamp connections; cortical hyphae often minutely spiculate (spicules less than 0.5 µm high, slender, sharp, cyanophilous), with more crystalline incrustation than medullary hyphae; ampulliform clamps overshot, tibiiform to aliiform, up to 14 µm broad, thick-walled (wall up to 1.2 µm thick), unornamented. Subicular hyphae identical to rhizomorph hyphae except: 1) little crystalline incrustation produced; 2) spiculate hyphae common throughout; and 3) additional hyphal type produced as branches of deep-seated subicular hyphae; these 1-2 µm diam, tightly coiled and/or with copious peg-like blunt branches, occasionaly terminating with flagellar, slender (less that 0.5 µm diam) tips. Tramal hyphae of upper branches 3-10 µm diam, hyaline, thick-walled (wall up to 0.5 µm thick), relatively straight, conspicuously clamped, parallel, tightly packed, not adherent or agglutinated. Subhymenium extensive, pseudoparenchymatous. Hymenium thickened significantly; basidia 40-50 X 7-9 µm, clavate, clamped; contents homogeneous at all ages; sterigmata 4, up to 7 µm long, slender, slightly incurved.
Basidiospores (n = 64/8) 8.6-12.6 X 2.9-4.0 µm (E = 2.36-3.78; Em = 3.02; Lm = 10.68 µm), elongate comma-shaped, elongate teardrop-shaped or boletoid, smooth, thin-walled, hyaline; contents usually homogeneous or with 1-several refringent guttules; hilar appendix curved, broad, not prominent.
Specimens examined: ARGENTINA, Prov. Misiones, Dpto. Frontera, date unknown, coll. Singer no. M1145 (FH); BHUTAN, TENN 44245; BOLIVIA, San Buena Vista, 1.IX.22, Mulford Biological Expedition of the Amazon Basin, coll. O.E. White, no. 1235 (NY); TENN 32201; BRAZIL, RHP 4437; Sao Paulo, Picado Lindly, Parque do Estado, 15.II.60, coll. M.E.P.K. Fidalgo, SP47228 (NY); CHINA, Yunnan Prov., Xishuanbanna, TENN 45675; CUBA, Nouvelle Sophie, 2.IX.?, Fungi Cubensis Wrightiana no. 235 (K, lectotype specimen of Clavaria cervina; isolectotype, FH); location unknown, 14.VIII.?, Fungi Cubensis Wrightiana no. 256 (lectotype specimen of Clavaria pallida; isolectotype, FH); JAMAICA, no location, no date, no collector, ex herb. Berkeley (K; type specimen of L. compressa); MALAYSIA, Pehang, Tembeling, 20.XI.30, Corner no. 551 (K); Malay Peninsula, South Ketch, Gna Ninik, 24.X.27, coll. M.R. Henderson, det. E.J.H. Corner, no. 19620 (K; TENN 31753); PHILIPPINES, location unknown, date unknown, coll. P.S. Cumming, no. 2042 (K; type of L. surculus); UGANDA, TENN 31476; UNITED STATES, Florida, TENN 38409; Puerto Rico, TENN 32270.
Commentary: Corner's (1950) discussion of L. surculus perfectly fits these specimens, even to the envelopment of the lower basidiome by subicular hyphae. Corner also mentioned the indefinite dimensions of the subiculum.
The group of specimens on which the above circumscription was based originated from Malaysia, tropical China, the Philippine Islands (if the type specimen of L. surculus is included for other than spore characters), tropical North America (southern Florida), eastern Bolivia, tropical Brazil and Uganda. In my opinion, then, L. surculus seems pantropical in distribution.
As noted by Corner (1950, 1970), basidiospores are strangely rare in the hymenium of herbarium specimens, although adequate spores are shed to give a spore print within a few hours. It is hazardous to base taxonomic identification on spore measurements when often not more than 5-6 spores can be observed after a long search in a single microscope mount. Moreover, young basidiospores are not only short, but significantly more slender than those at maturity. When such spores are inadvertantly included in spore statistics, Lm values decrease while Em values remain more or less constant or increase slightly. Nonetheless, after diligent search, it is usually possible to estimate spore length ranges, and when this is done, L. surculus and L. byssiseda are quite different. It is also clear, for example, that New Zealand specimens (Petersen, 1988; basidiospores 15-24 X 4.5-6 mm) do not represent the same taxon as the collections used for the circumscription above. Neither do Chinese specimens under L. byssiseda (Petersen, 1989; basidiospores 15.5-16.9 X 4.0-4.3 µm), collected in northern conifer forests.
The type specimen of L. surculus (Philippines, Cumming no. 2042, herb. Berkeley, K) was examined by Corner (1950) and by me, but no spores have been observed, so assignment of this name to all other specimens must be done on circumstantial evidence of basidiome and subiculum morphology.
For many years (most recently Petersen, 1988) I considered L. byssiseda and L. surculus to be so similar as to be inseparable taxonomically, taking my lead from Corner (1950). Nonetheless, I considered L. surculus to be a tropical and Southern Hemisphere organism, while L. byssiseda was considered to fruit across the North Temperate Zone. It now becomes obvious that the two taxa are separable by spore statistics, and that L. surculus is again divisible into several specific and infraspecific entities.
Lentaria surculus
variant A (Figs. 3, 4)
Some specimens collected in areas of northern and eastern South America and southern Mesoamerica bear spores somewhat stouter than those of typical L. surculus as described above. Only two specimens were accompanied by even rudimentary notes, so the range of basidiome colors is not accurately known. In addition, specimens from eastern Asia are similar, although probably coincidentally so. An abbreviated description can be offered.
Subiculum a felty mat involving slender rhizomorphs within the substratum, extensive to not so, off-white. Stipe base off-white to "cinnamon buff; 6B4." Branches and apices more delicately dissected than in other forms; lower and middle branches "cinnamon;" 6B5. Apices elongate awl-shaped,"pale pinkish cinnamon;" 6A2. Hymenium often unilateral, with off-white sterile surface decurrent from axils; axils lunate below, narrowly rounded above; branches in up to six ranks, terete to flattened below, terete above, sympodial to dichotomous throughout. Odor none. Taste distinctly bitter.

Commentary. Overall basidiospore measurements (n = 63/5; 10.8-15.8 X 3.6-5.8 µm; E = 2.36-3.70; Em = 2.90; Lm =13.45 µm) are somewhat larger than those reported above, and especially increased spore width reduces Em value somewhat. Nonetheless, other micromorphological characters fit those of typical specimens. Significant in light of the information below on other South and Central American collections, the subiculum in the eastern South American specimens is well-developed, with the usual extensive mat and inherent rhizomorphs. The wood substrate is invariably quite decorticate and loose, with copious mycelium and rhizomorphs. While it is possible that a cline may be found between basidiospore statistics in "variant A" and those of typical specimens, there currently exists a hiatus in spore length and width, although hardly reflected in E and Em values.
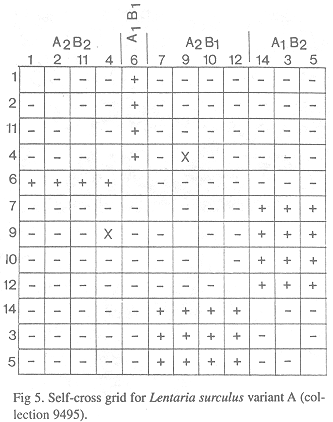
Mating system in "Variant A" (collection 9495, TENN 56523). When 12 SBIs were paired in all combinations, a tetrapolar mating system was revealed (Fig. 5). Isolate 6* = A1B1; 1, 2*, 4, 11 = A2B2; 3, 5, 14* = A1B2; 7*, 9, 10, 12 = A2B1 (asterisks mark isolates selected as tester strains and deposited at CBS). No barrage or flat reactions were noted, although some mating exhibited lethal reactions on AERIAL mycelium, not agar-surface. Subordinate mating types were assigned arbitrarily, therefore.
Culture morphology. Colonies white to very pale pearl gray, densely minutely wooly, more so approaching colony margin. Colony margin abruptly diffuse to rough or irregular.
Hyphal differentiation as follows: 1) aerial hyphae always showing crystal deposition, usually very lightly spiculate, but often with short lengths near agar surface heavily incrusted with rock-like crystals; 2) agar-surface (and some juxtaposed aerial) hyphae usually diverticulate, with diverticulae short, cog-like; and 2) common agar-surface hyphae (never aerials) with elongate-alliiform swellings juxtaposed to septa (always simple septa, never clamped). Clamp connections were often scarce and very local, best seen on aerial hyphae near the agar surface. Agar-surface hyphae rarely bore clamps. Clamps were usually seen at the crotches of diverticulate branches.
Specimens examined: ARGENTINA, TENN 31475; BRAZIL, Pernambuco, TENN 34873; TENN 32267; COSTA RICA, Prov. Puntarenas, Canton Coto Brus, Hacienda La Amistad, 8o 54.218' N, 82o 47.401' W, 4.VII.98, coll. RHP, on well-rotted sticks, no. 9495 (TENN 56523). PANAMA, Prov. Panama, Gatun Lake, Barro Colorado Island, Shannon Trail, 10.VIII.97, coll. C.L. Ovrebo, Ovrebo no. 3592 (OVREBO; TENN 56937); same location, AVA Trail, 8.VIII.97, coll. C.L. Ovrebo, Ovrebo no. 3571 (OVREBO; TENN 56936). VENEZUELA, TENN 47557; BHUTAN, TENN 44244; RUSSIA, Terr. Siberia, Dist. Tara, Omsk, 2.IX.28, coll Murashkinsky no. 015 (FH).
Lentaria surculus
variant B (Fig. 6)
From Colombia, Panama and tropical Mexico have come some specimens which differ from the pantropical and eastern South American collections in three significant ways: 1) subiculum greatly reduced, often to a small, tough pad only twice the diameter of the discrete basidioma; 2) only one basidioma arises from one subiculum; and 3) basidiopores are narrower and therefore Em value is higher. A description follows.
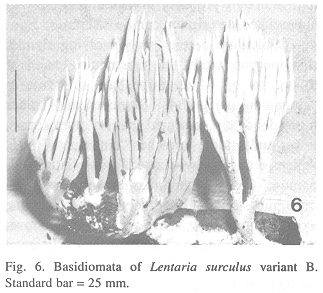
Basidiomata up to 10.5 X 5 cm, branched repeatedly, arbuscular. Stipe up to 3 X 1.2 cm, arising from a subicular mat, single or rarely cespitose, white, off-white or beige below, reluctantly bruising or decaying to 8E8, occasionally covered with mycelial mat similar to subiculum; subiculum limited to a small, tough, felty mat, or occasionally more widespread, thin-lacerate at margin, without inherent rhizomorphs, white, pallid brown where rubbed, up to 5 X 3 cm (usually more limited), up to 2 mm thick, densely felty, often subtended by discrete, ropy rhizomorphs up to 2 mm thick; stipe flesh white to off-white, punky to felty, drying light in weight, easily pierced. Branches in up to five ranks, terete above, flattened or lobed in cross-section below, erect to strict, avellanous tan ("fawn color," "mikado brown" below; 7C5-6) to ochraceous tan ("ochraceous tawny," "pinkish buff" upward; 6B-C6, 6A3); internodes remaining more or less constant in length except common dichotomous apices; axils rounded throughout, lunate apically. Apices dichotomous to prolonged awl-shaped, up to 1.5 cm long, pale tan to pallid buff, white to "pale cinnamon pink;" 5A2 at tips. Odor mild; taste bitter. Habit on hardwood logs and sticks, including Quercus.
Macrochemical reactions: FSW = olive green.
Hyphae of rhizomorphs 1.5-3.5 µm diam, hyaline, thick-walled (wall uniformly 0.5 µm thick), infrequently but conspicuously clamped; wall of medullary hyphae smooth, those of cortical and surface hyphae spiculate; spicules crowded, less than 0.5 µm long, extremely sharp, cyanophilous; ampulliform clamps up to 14 µm broad, overshot, thick-walled (wall up to 1 mm thick), without ornamentation. Stipe tramal hyphae 2-9 µm diam, hyaline, thick-walled (wall 0.5-1 µm thick), conspicuously clamped, loosely packed inward, interwoven, occasionally with narrow clampless hyphal tips (2-2.5 µm diam; up to 400 µm long) reminiscent of skeletal hyphae; ampulliform clamps up to 14 µm broad, overshot to tibiiform,. thick-walled (wall up to 1 µm thick), unornamented. Tramal hyphae of upper branches 2-9 µm diam, hyaline, thin- to thick-walled (wall commonly up to 0.5 µm thick), conspicuously clamped; ampulliform clamps common, up to 13 µm broad, overshot, not usually thick-walled, often with amorphous inclusions but not ornamented. Subhymenium extensive; hyphae 1.5-2.5 µm diam, hyaline, conspicuously clamped, highly branched, tightly packed. Hymenium thickened significantly; basidia 48-55 X 7-8.5 µm, clavate to narrowly sphaeropedunculate, clamped; contents multiguttulate to multigranular at maturity, the guttules/granules yellow-refringent; sterigmata 4, up to 8 µm long, slender, straight, spindly; filiform hyphal tips occasionally exerted from hymenium (TENN 43721; Desjardin 4773).
Basidiospores (n = 41/4) 9.4-13.3 X 2.5-3.6 µm (E = 2.89-5.00; Em = 3.63; Lm = 11.26 µm), cylindrical to elongate comma-shaped to sway-backed, with suprahilar swelling, hyaline, thin-walled; contents several- to multiguttulate, the guttules yellow-refringent; hilar appendix gradual, inconspicuous.
Specimens examined: COLOMBIA, Dpto. Del Valle, Mun. Zarzal, Corr. "la Paila," hac. El Medio, 31.V.90, coll. A.E. Franco-Molano, no. 52, in mixed forest on wood (TENN, CUVC); Halling 6085 (TENN, NY); Departamento unknown, El Medio, Desjardin 4773, 4774, 4775, (TENN); MEXICO, (TENN 43721); Est. Chiapas, vic. Allemania, Finca Irlanda, ecological preserve, 15° 10.385' N, 92o 20.185' W, 21.X.97, coll. Graciella, no. 5074 (TENN 55921).
Commentary: At this time, it would be premature to assign infraspecific ranks and epithets to variants A and B, for both are known imperfectly, and largely from herbarium material. Two hypotheses seem valid, however: 1) L. surculus represents a morphotaxonomic complex, unlike L. byssiseda, which appears relatively uniform over its entire range; and 2) although Corner (1950, but not 1970) and Petersen (1988) have opined that L. byssiseda and L. surculus showed few differences, in fact, spore statistics together with geographic distribution separate them.
Mating System
of Lentaria Byssisedo
With separation between L. surculus and L. byssiseda made possible by microscopic characters, the report below can be furnished with taxonomic confidence, and with L. surculus variant A, above, may represent the first report of a mating system in the Gomphaceae s.l. The specimen represented a "stag-horn" form, with lower parts "pinkish buff;" 6A3 to "pale pinkish cinnamon;" 6A2. Branch apices were prolonged, acerose, "cartridge buff." Odor mildly pleasant.
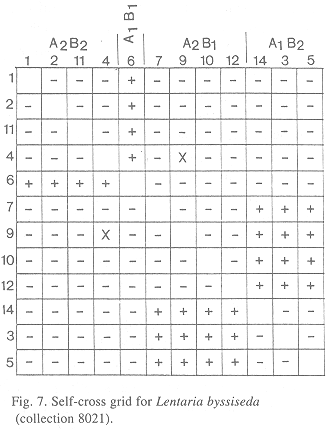
When 12 single-basidiospore isolates were paired in all combinations, a tetrapolar mating system was revealed (Fig. 7). Isolates 2*, 4, 6, 15 = A1B1; 10*, 13, 14 = A2B2; 19* = A1B2; 8*, 12, 16, 17 = A2B1 (asterisks mark isolates selected as tester strains and deposited at CBS). No discernible flat or barrage contact zone morphologies were detected, so subordinate mating types were assigned arbitrarily.
In many pairings, colonies were mutually repulsed (i.e. growth away from contact zone was much larger radius than into the contact zone), and even when isolates appeared to merge to the naked eye, they were really at different levels of submersion in the agar. Thus many pairings had minimal contact.
Culture morphology: Monokaryon colonies were white or off-white (except for 8021: 13, which was cream-colored), and consistently staurate (i.e. plumose, with plumes tendril-like and submerged). Colony odor was very weak, but perhaps salty or slightly penetrating. Dikaryon colonies produced off-white aerial mycelium as a thin haze. Dikaryon mycelium tended to overgrow the monokaryon donors, so that often there was a distinct dikaryon margin half way through one of the monokaryons.
Monokaryon cultures are characterized by the following hyphal types: 1) submerged, sclerified hyphal tips of generally acute-triangular shape, with sharp barb-shaped or awl-shaped side branches from a single major hypha; 2) aerial hyphae which act as rhizoids (i.e. arching through the air from one point to another), slender (3.5-5.5 µm diam), lightly incrusted with acicular crystals (quite like those in the rhizomorphs under fruitbodies of Lentaria), with clamp connections also lightly incrusted; and 3) very heavily incrusted hyphal tips which arise at the agar surface and extend for 100-400 mm into the air; crystals plate-like, appearing like miniature mica crystals. Because of all this hyphal differentiation, clamps were unusually difficult to discern.
With such large basidiospores, it might be expected that more than one nucleus would be included in each spore. Under epifluorescence microscopy only a few spores were observed, and nuclear number could not be estimated accurately. In two spores, however, at least two nuclei appeared. Even after culling isolates with clamp connections (three out of 25 putative single-basidiospore isolates), genetic evidence strongly indicates that extra-numerary nuclei are clonal, for if they had migrated into basidiospores randomly, a self-cross would show some hemicompatible combinations in which a single isolate mates with more than one mating type. Such phenomena were not observed. Moreover, with these results, it is most likely that after meiotic nuclei migrate into incipient basidiospores, they undergo one or more mitotic divisions.
Specimen used: UNITED STATES, North Carolina, Macon Co., Standing Indian Campground, 11.X.96, coll. RHP, on hardwood bough, no. 8021 (TENN no. 55580).
Interspecific pairings. When four single-basidiospore isolates of L. surculus (variant A: collection 9495, isolates 2, 6, 7 14) were paired with four single-basidiospore isolates of L. byssiseda (collection 8021, isolates 2, 8, 10, 19; n = 8), all pairings were interINcompatible, strongly indicating that the two collections represented two genetically dissimilar taxa. This was the expected result, since the collections had been identified morphologically as separate taxa.
Acknowledgements
I thank Clark Ovrebo and Roy Halling for the transparencies from which figures 3, 4 and 6 were made. Highlands Biological Station furnished laboratory facilities. Juan Luis Mata wrote the Spanish resumen. This project was supported, in part, by NSF grant DEB-9521526 (PEET Program).
Resumen
Se proponen los siguientes nombres para las especies de Lentaria nuevas para la ciencia: L. boletosporioides de Nueva Zelandia, L. caribbeana del Caribe y regiones vecinas, L. glaucosiccescens de Nueva Zelandia, y L rionegrensis de Argentina. Se propone a Lentaria javanica nom. nov., como nombre substituto para Clavaria compressa, un homónimo tardío de su nomenclatura. Se discute el agrupamiento infraespecífico dentro de L. surculus basado en estadísticas de las mediciones de las esporas. Se describen los sistemas reproductivos para L. surculus y L. byssiseda, y se encontró que éstas dos especies son sexualmente interincompatibles.
References
Corner, E.J.H. 1950. A monograph of Clavaria and allied genera. Ann. Bot. Mem. 1: 1- 740 [ Links ]
Corner, E.J.H. 1957. Some clavarias from Argentina. Darwiniana 11: 193-206. [ Links ]
Corner, E.J.H. 1970. Supplement to "A monograph of Clavaria and allied genera." Beih. Nova Hedwigia 33: 1-299. [ Links ]
Fries, E.M. 1821. Systema Myvcologicum. Vol. 1. Gryphswaldiae. 518 pp. [ Links ]
Gordon, S.A. & R.H. Petersen. 1991. Mating systems in Marasmius. Mycotaxon 41: 372- 386. [ Links ]
Kornerup, A. & J.H. Wanscher. 1967. Methuen handbook of colour. Second edition., London, Methuen. 243 pp + 30 clr. pls. [ Links ]
Petersen, R.H. 1967. Notes on clavarioid fungi. VII. Redefinition of the Clavaria vernalis C. mucida complex. Amer. Midl. Nat. 77: 205-221. [ Links ]
Petersen, R.H. 1971a. Interfamilial relationships in the clavarioid and cantharelloid fungi. pp. 345-374 IN "Evolution of the Higher Basidiomycetes, R.H. Petersen, ed., University of Tennesse, Knoxville, Tennessee. [ Links ]
Petersen, R.H. 1971b. Types studies in clavarioid fungi. IV. Specimens from herbarium Fries at Uppsala, with notes on cantharelloid species. Friesia 9: 369-388. [ Links ]
Petersen, R.H. 1972. Notes on clavarioid fungi. XII. Miscellaneous notes on Clavariadelphus, and a new segregate genus. Mycologia 64: 137-152. [ Links ]
Petersen, R.H. 1974a. Notes on clavarioid fungi. XIV. Cultures of Lentaria byssiseda. Mycologia 66: 530-532. [ Links ]
Petersen, R.H. 1974b. Contribution toward a monograph of Ramaria. II. Exclusion of Ramaria pinicola (Burt) Corner. J. Elisha Mitchell Sci. Soc. 90: 66-68. [ Links ]
Petersen, R.H. 1974c. Notes on clavarioid fungi: Three new species of Clavariadelphus. Trans. Brit. Mycol. Soc. 63: 469-474. [ Links ]
Petersen, R.H. 1985. Type studies in the clavarioid fungi IX. Miscellaneous taxa, with a section on Tremellodendropsis. Persoonia 12: 401-413. [ Links ]
Petersen, R.H. 1988. The Clavarioid Fungi of New Zealand. DSIR Bull. 236: 1-170. [ Links ]
Petersen, R.H. 1989. Some clavarioid fungi from northern China. Mycosystema 2: 159- 173. [ Links ]
Ridgway, R. 1912. Color standards and color nomenclature. Washington, D.C., publ. priv. 43 pp + 53 pls. [ Links ]
Singer, R. 1969. Mycoflora australis. Beih. Nova Hedwigia 29: 1-405. [ Links ]
1 Botany Department, University of Tennessee, Knoxville, Tennessee 37996-1100, USA. Fax: 1-423-974-0978, e-mail: repete@utk.edu

