










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.48 n.2-3 San José Jun. 2000
Colombia y diagnóstico del género
César Román-Valencia 1
Recibido 7-IX-2000. Corregido 13-III-2000. Aceptado 22-III-2000.
Abstract
Three new fish species are described: Bryconamericus dahli from the basins of the Patia and Mira rivers; B. ichoensis from Chaparraido Creek, the upper Atrato River basin, and B. galvisi from upper Putumayo River. Bryconamericus dahli can be distinguished from other Bryconamericus species by body depth and the larger head. B. dahli is similar to B. caucanus, but can be distinguished by the number of anal fin rays, head width and maxilla. Bryconamericus ichoensis can be distinguished by its small size, absence of a spot on the caudal peduncle, and generally 27 to 30 anal fin rays. B. ichoensis may be closely related to B. multiradiatus, but can be distinguished by the number of unbranched rays in the dorsal, pectoral and pelvic fins, the number of predorsal scales, body depth, etc. Bryconamericus galvisi, can be distinguished from other species of Bryconamericus by its single peduncle spot, high branched anal fin ray, the lateral line scales count, and elongated body. This species is similar to B. caucanus and can be distinguished by the number of teeth on the maxilla and by the number of vertebrae. The genus Bryconamericus is a natural and valid group, which is related to Hemibrycon. Moreover the Knodus and Eretmobrycon are synonym of Bryconamericus
Key words
New species, Characoids, Tropical fish, Bryconamericus.
Las especies del género Bryconamericus son predominantemente plateadas, con una mancha humeral y peduncular oscura y una banda lateral plateada en los ejemplares vivos, la cual se extiende hasta los radios medios caudales (Géry 1977) y por presentar una mancha morada en el opérculo en peces vivos (Bussing 1998). Son abundantes en pequeñas quebradas de tipo primario, secundario y en las orillas de los ríos de ancho promedio 40 m caracterizados por presentar oxígeno disuelto alto (promedio 8 ppm) y pH alrededor de la neutralidad, de tamaño pequeño (máximo 12 cm de longitud estándar), poseen una sola hilera de dientes en el dentario, dos hileras de dientes en el premaxilar con cuatro dientes en la serie interna (en la mayoría de las especies) que son más largos que la serie externa, el bajo número de dientes a lo largo del borde anterior del maxilar, la curva simple del borde de la mandíbula superior, el segundo y tercer suborbital en contacto con el preopérculo a lo largo de su borde anterior y ventral, la ausencia de una bolsa glandular sobre la aleta caudal en los machos (Román-Valencia 1998), el canal preopercular no sobrepasa el borde antero dorsal del opérculo, la ausencia de huesos nasal y supraorbitales, la apofisis ventral del rinoesfenoides es cartilaginoso, los canales sensoriales del pterótico y supraorbital sin interrupción, el ceratobranquial con una segunda fila de pequeños rastrillos branquiales, el primer radial proximal de la aleta dorsal sin quilla, de cuatro a seis supraneurales entre los músculos epaxiales anteriores a la aleta dorsal, el extremo inferior de estos huesos se proyectan en medio de las espinas neurales, la forma de estos huesos son diferentes a los radiales proximales de la aleta dorsal, por la ausencia de una apofisis anteromedial. La cintura pectoral presenta nueve radiales proximales, el extremo proximal del radial latero anterior tiene un proceso articular extensivo corto debajo de la superficie ventral de la escapula y el arco branquial en forma de z. Las especies de Bryconamericus aparecen distribuidos en una variedad de ecosistemas de agua dulce de las partes bajas y altas de Sur y Centro América a ambos lados de la cordillera de los Andes (Vari & Siebert 1990, Jiménez et al. 1998).
Los nuevos taxones aquí descritos se relacionarían con los pertenecientes a la parte transandina de Suramérica. Géry (1977) creó dos grupos artificiales en base a la longitud de la aleta anal y la cual está correlacionada con el número de escamas transversales. En este caso, B. ichoensis, B. galvisi y B. dahli se incluirían en el grupo peruanus.
Hasta el momento se han descrito 47 especies de Bryconamericus (Meek & Hildebrand 1916, Eigenmann 1927, Schultz 1944, Fowler 1943, 1944, 1945, Dahl 1960, Géry 1966, Dahl & Medem 1964, Vari & Siebert 1990, Bizerril & Araujo 1992, Malabarba & Kindel 1995, Román-Valencia 1998) que se extienden desde Argentina hasta Nicaragua; con mayor diversidad en la vertiente del Atlántico, en particular en la cuenca Amazónica (Vari & Siebert 1990). Para Colombia se registraron 18 especies (Cala & Román-Valencia 1994), distribuidas en todas las cuencas hidrográficas (Cala 1977, 1991, Román-Valencia 1988, 1990, 1993, 1995, Román-Valencia & Acero 1992).
En una investigación que adelanta el autor tendiente a la revisión del género para Centro America y el Norte de Suramerica, se observaron diferencias marcadas en unos ejemplares de Bryconamericus de la región trans y cis Andina de Colombia que determinaron el presente artículo.
Materiales y Métodos
Las medidas se tomaron con un calibrador digital hasta centésimas de mm y se expresaron como porcentajes de la longitud estándar y la longitud de la cabeza. En los recuentos de radios, escamas y dientes se utilizó estereoscopio, aguja de disección y coloreando en algunos casos con azul de metileno. Medidas y recuentos se realizaron sobre el lado izquierdo de los ejemplares, excepto cuándo éstos estaban deteriorados en tal lado. Recuentos y medidas siguen en general a Vari & Siebert (1990). El conteo de vértebras y radios procurrentes de la aleta caudal fueron obtenidos de radiografías y espécimenes clareados y teñidos de acuerdo al método de Taylor & Van Dyke (1985). Los ejemplares se compararon con material tipo depositado en la división de peces del Museo Nacional de Historia Natural, Instituto Smithsonian en Washington (USNM); en el Field Museum of Natural History, Chicago (FMNH) y en el Museo de Ciencias Naturales de Filadelfia (ANSP). El material se depositó en la unidad de Ictiología del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia, Bogotá (ICNMNH) y en el laboratorio de Ictiología, Departamento de Biología, Universidad del Quindio, Armenia, Colombia (IUQ). Excepto IUQ el código de las instituciones sigue a Leviton et al. (1985).
Se hicieron determinaciones diurnas, en las localidades típicas, de las siguientes variables: oxigeno disuelto y temperaturas superficiales del agua y del ambiente, determinado con oximetro. El pH, determinado con peachimetro portátil. Ancho y profundidad de la corriente de agua, medidos con un decámetro y una vareta graduada en centímetros. Tipo de sustrato y color del ambiente acuático, calificados por observación directa. Coordenadas y altura, con un Sistema de Posición Global (SGP) 4000XL.
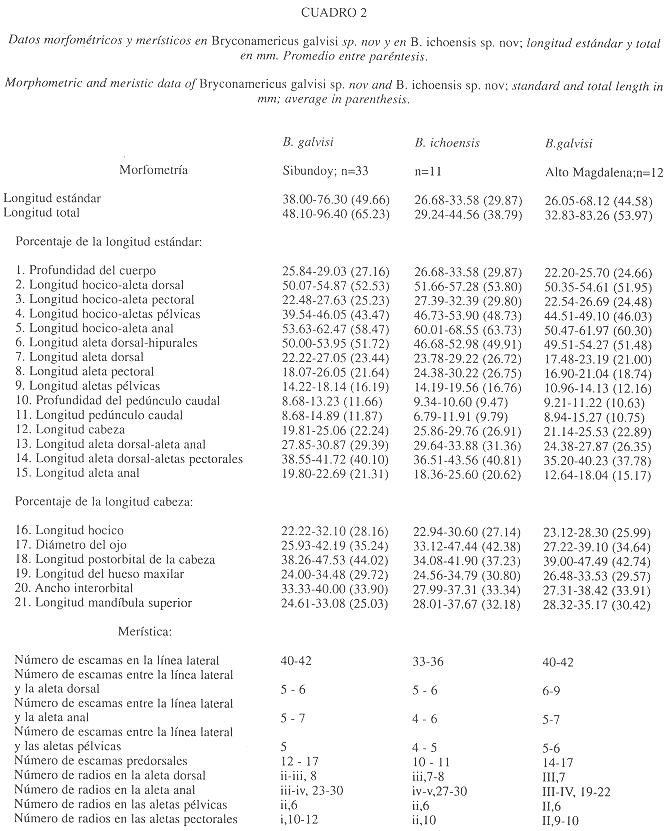
Los 21 caracteres morfometricos utilizados en este trabajo (Cuadros 1 y 2) fueron útiles para efectuar un análisis de componentes principales (ACP) y un análisis cluster o de clasificación. Además, se realizo un análisis de varianza para determinar significancias entre sexos de B. dahli y B. galvisi y entre los taxones aquí descritos. En estos casos se utilizo el paquete Spad.n integrado, versión 2.5 P.C. (Cisia 1994) y el Stat Graph.
Holotipo: ICNMNH 2722, hembra, 52.8 mm LS, Colombia, Departamento de Nariño, sistema del río Patia, cuenca del río Telembi, Quebrada Babosa afluente río Sabunde, en la vía a Tumaco Km. 98, a dos km. de la margen derecha (aprox. 78º 23 W y los 1º 37 N) ; G. Galvis & M. Camargo; marzo 11 de 1995.
Paratipos: siete espécimenes colectados con el holotipo, ICNMNH 2349; ICN-MHN 2339, 21 ejemplares, Colombia, Departamento de Nariño, cuenca del río Mira, quebrada Angostura a 200 m de la margen derecha vía a Tumaco Km. 89; G. Galvis; marzo 11 de 1995. ICNMNH 3450, seis espécimenes, Colombia, Departamento de Nariño, Municipio de Barbacoas, vereda Gualcala, cuenca del río Patia, Quebrada agua limpia en el camino de piedra verde a Gualcala, mas o menos 360 m; G. Arango, enero 31 de 1980. ICNMNH tres ejemplares, Colombia. Departamento de Cariño, cuenca río Pata, río Nambi; G.Galvis; marzo 1995. IUQ 218, 14 ejemplares, Colombia, Departamento de Nariño, Municipio de Tumaco, sistema del río Patia, Quebrada Babosa, afluente del río Sabunde, cuenca del río Telembi en la vía Km 98, a dos Km. margen derecha; C. Román-V. & J. Fernández; 11 de abril de 1998. ICNMNH 2353, cinco ejemplares, Colombia, Departamento de Nariño, Municipio de Tumaco , cuenca río Patia, Quebrada Pianurbi en la vía a Llorente cerca a Caunapi; G. Galvis; marzo 11 de 1995. IUQ 219, 13 ejemplares, Colombia, Departamento de Nariño, municipio de Tumaco, cuenca del río Patia, Quebrada Tortuguera en la vía Km. 98, margen derecha 50 m ; C. Román-V. & J. Fernández; 11 de abril de 1998. IUQ, 220 cuatro ejemplares, Colombia, Departamento de Nariño, municipio de Tumaco, cuenca del río Mira Quebrada Angostura en la vía Km. 98, 300 m margen izquierda; C. Román-V. & J. Fernández; 11 de abril de 1998.
Diagnosis: La especie se distingue por un cuerpo robusto relativamente profundo en ejemplares inmaduros y adultos, por la presencia de una mancha peduncular oscura que se prolonga sobre los radios medios caudales y la presencia de sólo tres dientes sobre el maxilar.
English Diagnosis: The species can be distinguished by the body depth, by the presence of a dark peduncular spot that extends in to median rays of the caudal fin, and by to having only three teeth on the maxilla.
Descripción: Cuerpo robusto y ligeramente comprimido en ejemplares menores de siete cm de largo estándar. Perfil dorsal de la cabeza en línea recta, distintamente convexo desde el labio superior a través de los narinas a nivel de la parte dorsal de la cabeza a la altura de las órbitas. Perfil dorsal del cuerpo curvo desde el borde posterior de la cabeza a nivel del supraoccipital hasta el origen de la aleta dorsal; en línea oblicua a nivel postero dorsal del cuerpo y posterior a la aleta adiposa a nivel del pedúnculo caudal. Perfil ventral del hocico hasta el origen de las pélvicas notablemente convexo, más pronunciado en la parte posterior de las aletas pectorales. La mayor profundidad del cuerpo localizada en la parte anterior del origen de la aleta dorsal. Pedúnculo caudal lateralmente comprimido en ejemplares menores de siete cm de longitud estándar, profundidad en machos adultos (11.07-12.64%) siendo menor en las hembras adultas (9.89-10.92%) y es significativa (análisis de varianza F=9.57; P= 0.0057). Dimorfismo sexual representado por la presencia en los machos de espinas en los radios de las aletas anal y pélvicas.
Cabeza relativamente larga; hocico corto. Mandíbulas iguales, boca terminal; labios blandos, flexibles que no cubren la hilera externa del premaxilar cuando la boca se halla cerrada; borde ventral de la mandíbula superior ligeramente cóncavo; el extremo posterior del maxilar se extiende debajo de la orbital, alcanza una línea trazada a través de 1/3 del ojo. Narinas sobre cada lado muy cercanas entre si; abertura de las narinas posteriores verticalmente ovoide; abertura de las narinas anteriores presenta una excrecencia membranosa, extendiéndose en el borde posterior, separando las dos narinas. Órbita del ojo libre, sin párpado.
Presenta seis huesos infraorbitales osificado con un segmento de canal latero sensorial. Segundo infraorbital mas largo y ancho, su borde ventral y posterior en contacto con la superficie lateral del preopérculo. Cuarto infraorbital sin ramificación posterior del canal latero sensorial. El segmento del canal latero sensorial en el sexto infraorbital consiste de un solo tubo. No presentan supraorbital.
Mandíbula inferior con una sola serie de cuatro grandes dientes sobre cada dentario, seguidos por seis pequeños dientes que disminuyen gradualmente a partir del quinto hasta el décimo diente. Los cinco primeros dientes típicamente tricúspides con una cúspide central mucho mayor. La segunda cúspide a menudo con una muy pequeña cúspide en el borde lateral. El sexto diente bicúspide y del siete al diez unicúspides y curvos.


Dos hileras de dientes sobre el premaxilar; seis a siete largos dientes de la primera hilera externa, orientados de manera alterna a un eje principal, o sea, en forma de zigzag. Hilera externa de dientes bicúspides, con una cúspide central mucho más larga. Cuatro dientes en la fila interna del premaxilar con cuatro a cinco cúspides, la central mucho mayor. Maxilar con tres dientes tricúspides ubicados en el extremo anterior de su borde.
Borde de la aleta dorsal redondeado, el primer radio simple y los dos primeros radios ramificados mas largos que los demás; los radios dorsales tanto en juveniles como en adultos no alcanzan el borde de la adiposa. Aleta adiposa pequeña en todos los espécimenes. Borde de las aletas pectorales convexo, sus extremos sobrepasan el origen de las aletas pélvicas. Borde de las aletas pélvicas redondeado; no alcanzan el origen de la aleta anal. Aleta caudal bifurcada, con lóbulos largos y terminan en punta o filamento nueve o diez radios caudales procurrentes tanto dorsal como ventralmente.
Escamas cicloideas. Aleta caudal sin escamas. Aleta anal con una o dos series de escamas que cubren las bases de los radios anales. 36-39 escamas con poros de la línea lateral se extienden desde el supracleitrom hasta la unión de los hipurales, en unión con la base de la aleta caudal. Poros de la línea lateral forman una curva desde la primera hasta la sexta escama, el resto en línea recta; dos a tres series de escamas se extienden mas halla de los hipurales, en unión con la base de la aleta caudal.
Número total de vértebras: 33-36
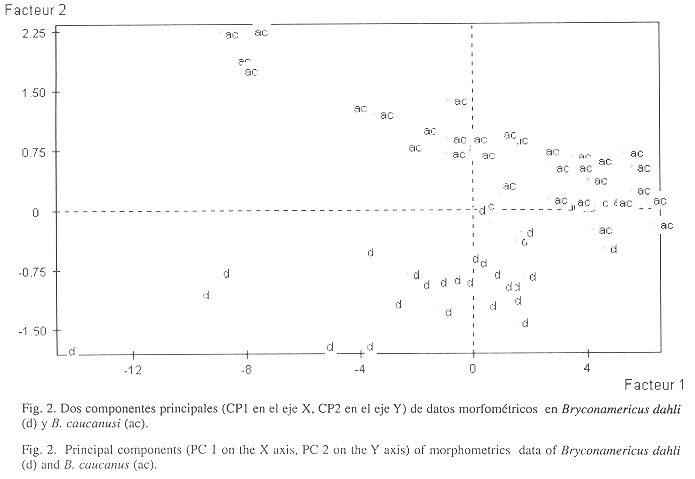
Comparaciones: B. dahli es muy similar a B. caucanus pero se diferencian en el número de radios de la aleta anal (31-35 en B. dahli, 25-28 en B. caucanus); el número de vértebras es mayor (37 en el nuevo taxon y 34 a 35 en B. caucanus); el hueso maxilar es menor que el diámetro del ojo en B. dahli, en B. caucanus el ojo es igual al maxilar; mientras que la longitud de la cabeza es mayor en la especie nueva al compararla con la longitud cabeza de B. caucanus. El analisis de componentes principales (Fig. 2) indicaron que estos taxones se distinguen en el eje 1 por la posicion de las aletas pelvicas con relacion al extremo del hocico y a la longitud de la aleta dorsal; en el eje 2, por el diametro del ojo, las longitudes postorbital y del pedunculo caudal y por la profundidad del cuerpo. La primera componente explica el 87.85% de la varianza total, y entre la primera y segunda componentes se explica el 92.09% de la variabilidad total. El nuevo taxon se aproxima a B. simus respecto al número de escamas en la línea lateral y en la longitud de la cabeza; se diferencian en el número de radios en la aleta anal (26-30 para B. simus, 31-34 en B. dahli), en la profundidad del cuerpo (3.4 para B. simus, 2.75-3.00 en B. dahli), en el ancho interorbital no es mayor que el diámetro del ojo para el nuevo taxon, el ojo es mayor en B. dahli al compararlo con el diámetro del ojo en B. simus.
B. dahli se diferencia de B. scopiferus (= B. emperador) igualmente registrada para la cuenca del río Mira en la profundidad del pedúnculo caudal, en la longitud de la cabeza y en el número de escamas predorsales. Además de lo anterior, B. dahli se distingue de B. miraensis, a en lo relacionado con la cabeza (más grande en la especie nueva), por el número menor de dientes grandes en el dentario (cuatro en B. dahli, ocho a nueve en B. miraensis), en el menor número de escamas predorsales (diez a 12 en sp nov. 14-17 en B. miraensis), en el mayor número de radios ramificados de la aleta dorsal (nueve a diez en B. Dahli, siete en B. miraensis), en el número mayor de dientes de la fila externa del premaxilar tres a cuatro en B. miraensis, seis en B. dahli) (cuadro 1), y por la ausencia de una banda lateral
Color en vivo: Área dorsal verde amarillenta, lateral y ventral blanco plateado el cual es mas pronunciado hacia la parte ventral del cuerpo. Aletas verde amarillentas con algunas manchas oscuras pequeñas. Mancha peduncular oscura prolongándose sobre los radios medios caudales; mancha humeral oscura muy difusa.
En todos los ejemplares se observó una mancha amarilla que abarca la región de las aletas pélvicas, anal, caudal, pectorales y caudal; la cual perdura aún por varios días en especímenes fijados en formol al 10% .
Distribución: Especie conocida de las cuencas de los ríos Patia y Mira.
Ecología: Los siguientes datos y observaciones fueron tomados en la localidad típica de la especie el día 11 de abril de 1998. Temperatura superficial del agua 26-27°C, oxigeno disuelto alrededor de los 9 mg/l, pH alrededor de la neutralidad, profundidad promedio 0.56 m, ancho promedio siete m, agua típicamente cristalina de sustrato conformado por piedra, arena y material orgánico en descomposición.
En los ejemplares capturados en la quebrada la Tortuguera, se logró observar que los machos expulsaron semen a la menor presión de sus flancos, a su vez las hembras expulsaban oocitos. Lo anterior sugiere que la especie lleva a cabo su evento reproductivo en sincronización con el inicio de las primeras lluvias (alrededor de marzo y abril).
En la quebrada Agua limpia se observó un sustrato conformado por grava y arena, de ancho aproximado 1.5-2.0 m.
Etimología: Epíteto especifico alusivo al Dr. George Dahl, por sus importantes contribuciones al estudio y conservación de los peces continentales y marinos de Colombia. Sus colecciones en el país fueron el inicio para la creación del museo mas grande de peces de agua dulce de nuestro país, en el Instituto de Ciencias Naturales, Universidad Nacional de Colombia en Bogotá (ICNMNH)
Holotipo: ICNMNH 2718 ; 31. 62 mm LE, Colombia, Departamento del Chocó, Municipio de San Francisco de Ichó, Alto Atrato, Quebrada Chaparraidó, afluente del río Ichó en la vía Tutunendo-San Francisco de Ichó, (aprox. 76º 38 W y los 5º 38 N); C. Román-V., Y. Palacios C. & C. Román-P. 20 dic. 1997.
Paratipos: diez espécimenes colectados con el holotipo, ICNMNH 2719
Diagnosis: La especie se diferencia por la ausencia de una mancha peduncular, en su caso presenta una banda oscura que se prolonga sobre los radios medios caudales, por presentar de 27-30 radios ramificados en la aleta anal y entre 33-36 escamas en la línea lateral.
English Diagnosis: The species can be distinguished by the absence of a peduncular spot, but there is a dark band on the midcaudal rays. There are between 27-30 branched anal fin rays, and between 33-36 along the lateral line scales.
Descripción: Cuerpo robusto en todos los espécimenes. Perfil dorsal de la cabeza en línea recta desde el hocico, la superficie dorsal de la cabeza convexo desde el borde del labio superior hasta una línea vertical trazada posteriormente a las narinas, levemente convexa desde esta línea hasta la espina del supraoccipital. Perfil dorsal del cuerpo ligeramente curvo desde el extremo posterior de la cabeza hasta el origen de la aleta dorsal, la superficie de esta región y hasta los últimos radios de la aleta dorsal hasta el pedúnculo caudal es convexo en todos los ejemplares. Perfil ventral del cuerpo generalmente curvo desde el extremo de la mandíbula inferior hasta el pedúnculo caudal. La máxima altura se localiza en el origen de la aleta dorsal. Pedúnculo caudal lateralmente comprimido. No se observó dimorfismo sexual en la especie.
Cabeza relativamente larga, perfil total redondeado anteriormente. Mandíbulas iguales, boca terminal; labios flexibles, con los extremos de los dientes cafés en la hilera externa, visibles cuando la boca esta cerrada, borde ventral de la mandíbula superior cóncavo; extremo posterior del maxilar se extiende debajo de la órbita, alcanza una vertical trazada en _ del diámetro del ojo; abertura de las narinas anteriores ovoide; mas ancho en el extremo anterior; excrecencia prolongándose en el borde posterior de la narina anterior, de tal manera que las separa. Borde orbital libre, sin párpado.
Seis infraorbitales osificado, cada uno posee un segmento de canal latero sensorial. Segundo infraorbital muy ancho, su borde ventral y posterior en contacto con la superficie lateral del preopérculo. No presenta supraorbitales.
Mandíbula inferior con una serie de ocho a nueve dientes, tricúspides con la cúspide central mas grande, de igual tamaño, seguidos por cuatro pequeños dientes tricúspides hacia los extremos del dentario .
Dos hileras de dientes sobre el premaxilar; dientes de la hilera interna más anchos. Cuatro a cinco dientes en la serie externa, orientados en zig zag. Fila externa de dientes bicúspides, con la cúspide central mucho mas larga. Cuatro dientes en la fila interna del premaxilar, con cinco cúspides, la cúspide central más larga. Maxilar con dos o tres pequeños dientes tricúspides localizados en el borde anterior; el diente dorsal del maxilar ubicado próximo al diente lateral del premaxilar.
Borde de la aleta dorsal redondeada, el segundo radio simple y el primer radio ramificado de diferente longitud; aleta dorsal deprimida no alcanza el borde anterior de la aleta adiposa en todos los espécimenes. Aleta adiposa moderadamente desarrollada en todos los ejemplares. Borde de las aletas pectorales redondeado, sus extremos alcanzan el origen de las aletas pélvicas en todos los espécimenes. Borde de las aletas pélvicas redondeado, sus extremos no alcanzan el origen de la aleta anal en todos los ejemplares. Aleta caudal bifurcada, lóbulos punteados; ocho radios procurrentes tanto dorsal como ventralmente.
Escamas cicloides. Aleta caudal sin escamas. Aleta anal con una serie de escamas que cubren las bases de los radios en toda su longitud. Escamas de la línea lateral se extienden desde el supracleitrom hasta los hipurales; las 13 primeras escamas de la línea lateral forman una curva, posteriormente en línea recta.
Número total de vértebras: 32-34
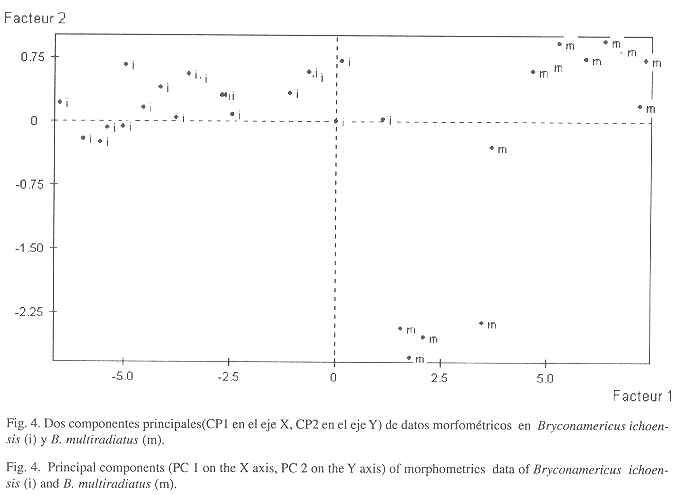
Comparaciones: B. ichoensis posiblemente está relacionada con B. multiradiatus debido a su reducido tamaño, su coloración en vivo y por que se aleja de las demás especies de Bryconamericus de Colombia. Se diferencian en el número de radios simples de la aletas dorsal (tres en B. ichoensis, dos en B. multiradiatus), pectorales (dos en el nuevo taxon, uno en B. multiradiatus) y pélvicas (dos en el nuevo taxon, uno en B. multiradiatus), en el número menor de escamas predorsales (10-11 en el nuevo taxon, 16 en B. multiradiatus), en la profundidad del cuerpo (29.87% en el nuevo taxon, 28.49% en B. multiradiatus), en la longitud hocíco-aleta dorsal (53.80% en el nuevo taxon, 48.10% en B. multiradiatus), en la longitud hocico- aleta anal (63.73% en el nuevo taxon, 54.02% en B. multiradiatus), en la longitud hocico-aleta pectorales (26.75% en el nuevo taxon, 25.90% en B. multiradiatus), en la longitud del hocico-aletas pélvicas (16.76% en el nuevo taxon, 44.77% en B. multiradiatus), en la longitud del pedúnculo caudal (9.79% en el nuevo taxon, 14.80% en B. multiradiatus).

El análisis de componentes principales (Fig. 4) indicó que estos taxones se distinguen en el eje 1 por la longitud de la cabeza y por la posición de la aleta dorsal con relación al orígen de la aleta anal; en el eje 2, por la longitud de la aleta dorsal, por la posición de las aletas pectorales con relación al extremo del hocico y por la profundidad del pedúnculo caudal. La primera componente explicó el 89.80% de la varianza total, y entre la primera y la segunda componentes se explicó el 94.56% de la variabilidad total. B. ichoensis se distingue de B.emperador, la especie afín que comparte su hábitat por el menor número de escamas en la línea lateral (cuadro 2), por no presentar mancha peduncular y por alcanzar un tamaño mucho menor (alrededor de los 30 mm de longitud estándar).
Color en vivo: Área dorsal amarilla; lateral y ventral blanca plateada la cual es más pronunciada en la parte ventral; aletas dorsal, pectorales, ventrales, anal amarillas. Una mancha peduncular oscura que se prolonga sobre los radios medios caudales. No presenta mancha humeral.
Distribución: Especie colectada solamente en la quebrada Chaparraidó en el sistema del río Ichó, cuenca del Alto Atrato, donde convive de manera simpátrica con B. emperador.
Ecología: Los siguientes datos y observaciones fueron obtenidos en la localidad típica de la especie, los días 20 y 23 de diciembre de 1997 entre las 13:00 hr y las 15:40 hr. Temperatura superficial del agua promedio 27.6ºC (26.5-27.2ºC), oxigeno disuelto 8.3 ppm (8-8.5 ppm), pH neutro, color del agua típicamente cristalina, con sustrato conformado por piedra, arena y material vegetal en descomposición, de ancho 5.7 m (4.5-7.2 m), profundidad 0.5 m (0.40-0.57 m). La especie se observó en un ambiente lótico y nadando contra la corriente en simpatria con B. ortholepis.
Etimología: Epíteto específico alusivo al sistema del río Ichó que pertenece el pequeño drenaje hábitat del nuevo taxon.
Holotipo: ICNMNH 2720, macho, 70.3 mm L.E., Colombia, Departamento del Putumayo, municipio de Sibundoy, cuenca del Alto Río Putumayo, Valle de Sibundoy (aprox. 77°23 W y los 5°00 N); O. J. Rodríguez; 13 julio 1993.
Paratipos: doce ejemplares colectados con el holotipo, ICNMNH 2721. IUQ 221, diez ejemplares colectados con el holotipo, ICNMNH 2721. IUQ 221, diez ejemplares Colombia, Departamento del Putumayo, municipio de Sibundoy, vereda Fátima, Quebrada Fátima sobre la vía a Orito, dos Km. adelante de Sibundoy; C. Román-Valencia, J. Tovar & H.Hoyos; junio 26 de 1998. IUQ 222, cinco ejemplares, Departamento del Putumayo, municipio de Sibundoy, vereda San Pedro, cuenca alta del río Putumayo, Río San Pedro, en la vía a Sibundoy un km. antes de la cabecera municipal; C. Román-V., J. Tovar & H. Hoyos; junio 27 de 1998. IUQ 223, cinco ejemplares, Colombia, Departamento de Putumayo, Municipio de Sibundoy, cuenca alta del río Putumayo, pequeño drenaje afluente del río Sibundoy, 100 m antes de llegar a la cabecera municipal sobre la vía a Orito; C. Román-V. J. Tovar & H. Hoyos; junio 26 de 1998. IUQ 310, tres ejemplares, Colombia, Departamento de Putumayo, cuenca alta del río Putumayo, quebrada el Cedro afluente del rio Putumayo , 200 m margen derecha en la via a Sibundoy; C. Román-V., J. Tovar & H. Hoyos; junio 27 de 1998.
Diagnosis: La especie se distingue dentro del género sobre las bases de los siguientes caracteres: carece de una mancha peduncular, por el número mayor de radios ramificados de la aleta anal (23-30) y de escamas en la línea lateral (40-42) y por el cuerpo relativamente elongado (27.16%) (Cuadro 2).
English Diagnosis: The species can be distinguished by the absence of a peduncular spot, by the great number of branched anal fin rays (27-30), the number of lateral line scales (40-42), and the elongated body (27.16%) (table 2).
Descripción: Cuerpo elongado en todos los ejemplares. Perfil dorsal de la cabeza en línea recta desde el hocico, su área superficial es convexa desde el borde del labio superior hasta una línea vertical trazada posteriormente a las narinas, cóncavo desde esta línea hasta la espina del supraoccipital. Perfil dorsal del cuerpo curvo desde el extremo posterior de la cabeza hasta el orígen de la aleta dorsal. Su área superficial generalmente convexa desde la base de los últimos radios de la aleta dorsal hasta el pedúnculo caudal. Perfil ventral del cuerpo ligeramente curvo desde el extremo de la mandíbula inferior hasta el pedúnculo caudal. La máxima altura se localiza en el orígen de la aleta dorsal. Pedúnculo caudal ligeramente comprimido, profundidad en los machos (19.81-22.3%) mayor en las hembras adultas (22.57-23.68%) y es significativa (análisis de varianza F=11.15; p=0.00074). Además, la longitud postorbital de la cabeza (45.95-47.53%) es menor en las hembras (42.11-45.56%) y es significativamente diferente (análisis de varianza F=12.772; P=0.0015); de igual manera la longitud de la cabeza (19.81-21.23%) y la distancia hocico-aleta pectoral (22.48-23.85%) es menor que la de las hembras (22.33-23.68 % la longitud de la cabeza y 25.80-27.63% la distancia hocico-aleta pectoral).y son altamente significativas (análisis de varianza F= 7.26; P=0.127 para la longitud de la cabeza y F=8.552; P=0.0074 para la distancia hocico-aletas pectorales).
Dimorfismo sexual representado por espinas en los radios de las aletas anal y pélvicas. Cabeza relativamente corta, y redondeada anteriormente. Mandíbulas iguales, boca terminal, labios flexibles, con los extremos de los dientes cafés más pronunciados en las hileras de los premaxilares; borde ventral de la mandíbula superior cóncavo; extremo posterior del maxilar se extiende hasta el borde de la órbita; abertura de las narinas anteriores redonda; excrecencia prolongándose hacia el borde posterior de la narina anterior, de tal manera que los separa. Borde orbital libre sin párpado.
Cuatro infraorbitales osificados, que no poseen un segmento de canal latero sensorial. Segundo infraorbital muy ancho, su borde ventral y posterior en contacto con la superficie lateral del preopérculo. No presenta supraorbitales. Mandíbula inferior con una serie de ocho a nueve dientes tricúspides, la cúspide central más grande, seguidos por siete pequeños dientes tricúspides hacia los extremos del dentario.
Dos hileras de dientes sobre el premaxilar, dientes de la hilera interna mas anchos. Cuatro dientes en la serie externa, orientados uno detrás del otro o en fila recta. Cuatro dientes en la fila interna del premaxilar, con cuatro cúspides, la central mas larga. Maxilar con dos a seis dientes pequeños tricúspides localizados en el borde anterior, el diente dorsal del maxilar ubicado próximo al diente lateral del premaxilar. Borde de la aleta dorsal truncada, sin embargo, el primer radio simple y el primero ramificado de diferente longitud; la aleta dorsal deprimida no alcanza el borde anterior de la aleta adiposa en todos los espécimenes. Aleta adiposa desarrollada en todos los ejemplares. Borde de las aletas pectorales redondeada, sus extremos apenas alcanzan el orígen de las aletas pélvicas en todos los individuos. Borde de las aletas pélvicas redondeadas, sus extremos alcanzan el orígen de la aleta anal. Aleta caudal bifurcada, lóbulos punteados, ocho radios procurrentes tanto dorsal como ventralmente. Escamas cicloideas. Aleta caudal sin escamas, excepto en su base que presenta tres pequeñas hileras. Aleta anal con una o dos series de escamas que cubren la base de los radios. Poros de la línea lateral en línea recta desde el supracleitrom hasta los hipurales en todos los ejemplares
Número total de vértebras: 37
Comparaciones: B. galvisi es muy parecida y posiblemente esta relacionada con B. caucanus, sin embargo se puede distinguir por el número de dientes en el maxilar (mas de tres dientes en el nuevo taxon, dos o tres dientes en B. caucanus), por el número de vértebras (37 en el nuevo taxon, 34-35 en B. caucanus) y por la longitud maxilar con respecto al diámetro del ojo (menor en el nuevo taxon, mientras que en B. caucanus es igual).
B. galvisi y B. caucanus son genéticamente distintas pero estrechamente relacionadas. Además, se pueden incluir en un grupo de Bryconamericus que incluye especies del Amazonas en Perú, Paraná en Argentina y cuenca del Maracaibo en Venezuela. Son completamente distintas de Bryconamericus de Centro America (Román-Valencia, en preparación)
B. galvisi se distingue de B. orteguasae y de B. hypopterus especies registradas para la Amazonia Colombiana (Fowler 1945) por la cabeza más pequeña, el hocíco más grande, por el diámetro del ojo ligeramente mas grande que la longitud del hocico, posee cuatro dientes a cada lado en la serie externa del premaxilar, y por presentar mas de dos dientes en el maxilar, por registrar siete pequeños dientes laterales adyacentes a la serie de ocho a nueve grandes dientes tricúspides en cada dentario y por registrar entre 40-42 escamas en la línea lateral que se ordenan en línea recta desde el supracleitrom hasta los hipurales.
Color en vivo: Area dorsal verde-oscuro; lateral y ventral blanco-plateado, más pronunciado en la parte ventral. Además se observó una banda lateral verde-amarillenta. Una mancha humeral oscura verticalmente elongada. Una ligera banda peduncular oscura, la cual se prolonga sobre los radios medios caudales. Aletas dorsal y caudal amarillo-claro; aletas pectorales y pélvicas blanco-plateada, en algunos ejemplares se observaron de un color rosado-claro.
Distribución: Valle del Sibundoy en alto río Putumayo.
Ecología: Los siguientes datos y observaciones fueron obtenidos en la localidad típica de la especie a una altura de 2060 m s n m , el día 26 y 27 de junio de 1998 a las 5:30 p.m. y entre las 8 a.m. y las 10 a.m. respectivamente. Temperatura superficial del agua promedio 13.8 °C (11.5-15.7°C), temperatura del aire promedio 15.2 °C (12-19.5°C), oxígeno disuelto 7.83 (7.3-8.8 ppm) ppm , pH 6.5, color del agua café, posiblemente a causa de la lluvias que azotaban la región en ese momento, al día siguiente se observo cristalina, con sustrato conformado por piedra y arena, de ancho 4.4 m (aunque también se logro capturar en la orilla de afluentes mayores (alrededor de 40 m de ancho), profundidad 0.68 m. La especie se observó en un ambiente lentico y nadando en aguas abiertas, pero al menor peligro se refugiaron en la vegetación ribereña. La mayoría de las quebradas donde habita B. galvisi, se halla rodeada por pasto para la cría y levante de ganado y en algunos casos sus riberas se hallan conformadas por vegetación de tipo arbustivo.
Etimología: Epíteto especifico alusivo al biólogo Germán Galvis, profesor del Depto. de Biología de la Universidad Nacional, Santafé de Bogotá.
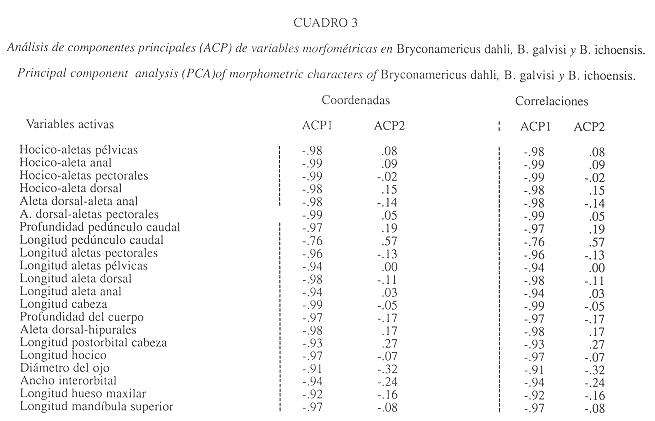
Comparaciones entre los taxones: Las especies se pueden diferenciar entre si, por los siguientes caracteres: ausencia de una mancha humeral en B. dahli; la línea lateral en su parte anterior no forma una curva, de cuerpo elongado, de cabeza pequeña y la longitud postorbital mayor en B. galvisi; la aleta dorsal presenta tres radios simples en B. ichoensis (cuadros 1 y 2). El análisis de componentes principales (Cuadro 3 y Fig. 6) indicaron que estos taxones se distinguen en el eje 1 por la posición de las aletas pectorales (análisis de varianza F=8.97; P=0.05), anal (análisis de varianza F=43.32; P=0.05) y la longitud de la cabeza (análisis de varianza F=15.45; P=0.05); en el eje 2, por el diámetro del ojo (análisis de varianza F= 22.15; P=0.05), el ancho interorbital (análisis de varianza F= 17.44; P=0.05), la longitud del maxilar (análisis de varianza F=10.63; p=0.05) y en la profundidad del cuerpo (análisis de varianza F=20.25; P=0.05). La primera componente explico el 91.34% de la varianza total, y entre el primero y segundo componentes se explico el 95.09% de la variabilidad total.

Discusión
El género Bryconamericus lo crearon Eigenmann et al. (1907), separándolo de Argopleura y Phenacobrycon por la ausencia de una bolsa de escamas en la base de los radios caudales, además, Eigenmann et al. (1907) anota su similitud con Astyanax, Mimagoniates, Hemibrycon y Piabina. Eigenmann (1922, 1927), Fowler (1945) y Weitzman & Fink (1985) plantearon que las especies descritas por Eigenmann (1913) tales como: Bryconamericus conventus, B. diquensis, B. chocoensis y B. magdalenensis son especies pertenecientes al género Argopleura. Los únicos caracteres que Argopleura y Bryconamericus comparten son plesiomórficos para su nivel taxonómico: se hallan en algunos otros taxones tetragonopterinos y por lo tanto no suministran información acerca de sus relaciones filogenéticas (Weitzman & Fink 1985).
Además, Eigenmann (1922; 1927) planteo que Bryconamericus boquiae, B. decurrens, B. dentatus, B. tolimae pertenecen al género Hemibrycon. El autor del presente trabajo logró examinar ejemplares de la localidad típica de Hemibrycon boquiae (especie endémica de la quebrada Boquia un pequeño drenaje del río Quindio, vereda Boquia, Salento, Quindío en Alto Cauca) y observó que convive en simpatria con B. caucanus. Las poblaciones de ambas especies en la quebrada Boquia se pueden separar por el número de dientes en el maxilar (ocho a nueve en H. boquiae, tres a cuatro en B. caucanus) y por el número de vértebras (41 en H. Boquiae, 34-35 en B. caucanus) .
Lucena (1993) reportó diez caracteres que podrian separar estos dos taxones, el correspondiente al maxilar observó que en Bryconamericus la extensión con dientes es menor o igual a la parte sin dientes (estado 1 de su análisis filogenético), mientras en Hemibrycon la extensión con dientes es mayor que el espacio sin dientes (estado 2). Por lo tanto, en el presente trabajo se considera a Bryconamericus diferenciable de Hemibrycon. De tal manera, válidas las especies H. Boquiae, H decurrens, H. dentatus y H. tolimae.
Por su parte, Schultz (1944) planteó que Knodus podría ser citado como un sinónimo de Bryconamericus basándose en la variabilidad de las escamas que cubren el lóbulo caudal en Knodus. Lo cual se comprobó por parte del autor del presente trabajo al examinar material de Venezuela, depositado en el Museo de Historia Natural La Salle, Caracas (MHNLS) y en el Museo de Biología del Instituto de Zoología Tropical, Universidad Central de Venezuela, Caracas (MBUCV) y determinados como Knodus. Por consiguiente es muy probable que muchas especies de Knodus se clasificaron dentro de Bryconamericus, y viceversa (Géry 1966). Además, Lucena (1993) no incluyó este grupo en su analisis filogenético. Aquí se considera a Knodus un sinónimo de Bryconamericus.
Fink (1976) describió al género Eretmobrycon, distinguiéndolo de Bryconamericus principalmente por el singular dimorfismo en el primer radio de la aleta caudal y en la forma de las espinas de los radios de las aletas anal y pélvicas. Sin embargo, el análisis de componentes principales sobre 21 caracteres morfometricos llevado a cabo en material de la localidad típica demostró que existe una relación estrecha de la forma y el tamaño entre el taxon descrito por Fink (1976) y Bryconamericus emperador. Distinguiéndose por el número de radios en las aletas pélvicas (dos radios simples y seis a siete ramificados en B. emperador, un radio simple y siete a ocho ramificados en B. bayano), por la ausencia en B. emperador de modificación alguna en los radios del lóbulo inferior de la aleta caudal, por el tamaño del hueso pélvico (en B. emperador es largo o sea de 4.20 mm de largo en promedio, en B. bayano es corto o sea 3.70 mm de largo en promedio), en el número de suborbitales (cinco en B. emperador, seis en B. bayano), en el número de branquias del primer arco branquial (9/11 en B. emperador, 8/12 en B. bayano), en el número total de vértebras (37-38 en B. emperador, 35-36 en B. bayano) En el presente trabajo se consideró válido el nombre Bryconamericus bayano y el género Eretmobrycon un sinónimo de Bryconamericus. La especie es endémica de la cuenca del río Bayano en la vertiente Pacifico de Panamá.
Agradecimientos
Este trabajo se llevó a cabo gracias a la financiación de la Universidad del Quindio (Depto. de Biología), la Fundación para la Promoción de la Investigación y la Tecnología del Banco de la República y de Colciencias. Jhon D. Lynch (ICNMNH), Germán Galvis (ICNMNH), Francisco Provenzano (MBUCV) y Anabel Perdices (STRI) leyeron el trabajo y plantearon valiosas sugerencias. Las siguientes personas e Instituciones facilitaron en préstamo y donación material de comparación: Eldredge Bermingham (STRI), William Bussing (UCR), Germán Galvis e Ivan Mojica (ICNMNH), E.B. Bohlke y Dominique D. Dagit (ANSP), Richard P. Vari y Susan L. Jewett (USNM) , Mary Anne Rogers y B. Chernoff (FMNH).
Resumen
Se describen tres nuevas especies: Bryconamericus dahli que habita en la cuenca de los Ríos Patia y Mira; B. ichoensis capturada en la Quebrada Chaparraidó, alto Río Atrato y B. galvisi colectada en el alto Río Putumayo. Bryconamericus. dahli se distingue de las especies conocidas de Bryconamericus por presentar un cuerpo relativamente profundo en ejemplares inmaduros y adultos, y por la cabeza más grande; es similar a B. caucanus, distinguiéndose en el número de radios de la aleta anal, en el tamaño de la cabeza y del maxilar. B. ichoensis puede diferenciarse de las especies conocidas por su pequeño tamaño (excepto con B. multiradiatus), por la ausencia de una mancha peduncular, sustituida por una banda oscura que se prolonga sobre los radios medios caudales, por 27-30 radios ramificados de la aleta anal; posiblemente se relaciona con B. multiradiatus por su enanismo, se distingue por el número de radios simples de las aletas dorsal, pectorales y pélvicas, por el número de escamas predorsales, por la profundidad del cuerpo. Bryconamericus galvisi, se puede distinguir dentro de Bryconamericus por una mancha peduncular, por el número mayor de radios ramificados de la aleta anal, por el número alto de escamas en la línea lateral y por el cuerpo elongado. Esta especie es muy semejante y relacionada a B. caucanus, distinguiéndose por el número de dientes en el maxilar, por la longitud del maxilar con relación al diámetro del ojo y por el número de vértebras. Se pueden diferenciar entre sí por la ausencia de una mancha humeral en B.dahli; la parte anterior de la línea lateral no forma una curva, de cuerpo elongado, de cabeza pequeña y la longitud postorbital mayor en B. galvisi; la aleta dorsal presenta tres radios simples en B. ichoensis. Se incluyeron datos del hábitat típico de los nuevos taxones. Se planteo que Bryconamericus es un grupo valido y natural, muy relacionado con el género Hemibrycon, mientras los géneros Knodus y Eretmobrycon son sinónimos de Bryconamericus.
Referencias
Bussing, A.W. 1998. Peces de las aguas continentales de Costa Rica. Segunda edición. Rev. Biol. Trop. 46: 468 p. (en Español e Ingles). [ Links ]
Bizerri, F. C.R S. & P.M.C. Araujo. 1992. Description a une nouvene espece au genre Bryconamericus (Characidae, Tetragonopterinae) du Brésil oriental. Rev. fr. Aquariol. 19: 65-68. [ Links ]
Cala, P. 1977. Los peces de la Orinoquia Colombiana. Lista preliminar anotada. Lozanía (Acta Zool. Col.) (24): 1-21. [ Links ]
Cala, P. 1991. Nuevos registros de peces para la Orinoquia Colombiana. I. Los rajiformes, clupeiformes, characiformes y gymnotiformes. Rev. Unellez Cienc. Tecnol. 4: 89-90. [ Links ]
Cala, P. & C. Román-Valencia. 1994. Lista y distribución geográfica de las especies de la familia Characidae (Pisces, Characiformes) en Colombia. Biol. Edu. 4:15-23. [ Links ]
Cisia. 1994. Oferta estudiante SPAD.N integrado, versión 2.5 P.C. CISIA, Francia [ Links ]
Dahl, G. 1960. New fresh-water fishes from western Colombia. Caldasia 8: 451-484. [ Links ]
Dahl, G. & F. Medem. 1964. Informes sobre la fauna acuática del río Sinú. CVM, Montería. [ Links ]
Eigenmann, C.H., W.L. Mc Atee & D.P. Ward. 1907. On further collection of fishes from Paraguay. Ann. Carniege Mus. 4: 139-140. [ Links ]
Eigenmann, C.H. 1913. Some results from an ichthyological reconnaissance of Colombia, South America. Part II. Indiana Univ. Stud. (131): 1-31. [ Links ]
Eigenmann, C.H. 1922. The fishes of the northwestern South America. Mem. Comp. Mus. (1): 1-348. [ Links ]
Eigenmann, C.H. 1927. The American Characidae. Mem. Mus. Comp. Zool. XLIII : 311-358; 417-428. [ Links ]
Fink, LW. 1976. A new genus and species of characid fish from the Bayano river basin, Panama (Pisces, Cypriniformes). Proc. Biol. Soc. Wash. 88(30): 331-344. [ Links ]
Fowler, W.H. 1943. A collection of fresh-water fishes from Colombia, obtained chiefly by brother Niceforo Maria. Proc. Acad. Nat. Sci. Phil. XCV:223-266. [ Links ]
Fowler, W.H. 1944. Fresh-water fishes from northwestern Colombia. Proc. Acad. Nat. Sci. Phil. XCVI: 227-248. [ Links ]
Fowler, W.H. 1945. Colombian Zoological Survey. Part I.- The fresh-water fishes obtained in 1945. Proc. Acad. Nat. Sci. Phil. XCVII: 93-135. [ Links ]
Géry, J. 1966. A review of certain tetragonopterinae (Characoidei), with the description of two new genera. The Aquarium J. 37 : 211-236. [ Links ]
Géry, J. 1977. Characoids of the world. T.F.H. Publ. Neptune 672 p. [ Links ]
Jiménez, J.L., Román-Valencia,C. & M. Cardona. 1998. Distribución y constancia de las comunidades de peces en la quebrada San Pablo, cuenca del río la Paila, Alto Cauca, Colombia. Actual. Biol. 20:21-27. [ Links ]
Leviton, E.A., R.H. Gibbs,Jr., E. Heal & C.E. Dawson. 1985. Standards in herpetology and Ichthyology: Part I. Standard Symbolic Codes for Institution Resource Collections in Herpetology and Ichthyology. Copeia, 1985:802-832. [ Links ]
Lucena, C.A.S. 1993. Estudo Filogenético da Familia Characidae com uma discussâo dos grupos naturais propostos (Teleostei:Ostariophysi:Characidae). Tesis Ph.D., Universidad de Sao Paulo, ix + 158 pp. [ Links ]
Malabarba, L.R. & A. Kindel. 1995. A new species of the genus Bryconamericus Eigenmann, 1907 from southern Brazil (Ostariophysi:Characidae). Proc. Biol. Soc. Wash. 108: 679-686. [ Links ]
Meek, E.S. & S.F. Hildebrand. 1916. The Fishes of the Freshwaters of Panama. Field Mus. Nat. Hist. (Zool.) X:217-374. [ Links ]
Román-Valencia, C. 1988. Clave taxonómica para la determinación de peces nativos de Quindio, subsistema alto rio Cauca, Colombia. Actual. Biol. 17: 107-114. [ Links ]
Román-Valencia, C. 1990. Lista y distribución de peces en la cuenca media del río Atrato, Chocó, Colombia. Caldasia 16:201-209. [ Links ]
Román-Valencia, C. 1993. Composición y estructura de las comunidades de peces de la cuenca del rio La Vieja, Alto Cauca, Colombia. Biol. Educ. (5): 8-19. [ Links ]
Román-Valencia, C. 1995. Lista anotada de los peces de la cuenca del río la Vieja, Alto Cauca, Colombia. Bol. Ecotropica (29):11-20. [ Links ]
Román-Valencia, C. 1998. Descripción de una nueva especie de Bryconamericus (Characiformes, Characidae) para la cuenca alta de los ríos Ariari y Meta, Colombia. Actual. Biol. 20: 109-114. [ Links ]
Román-Valencia, C. & A. Acero. 1992. Notas sobre las comunidades de peces del norte de Antioquia (Colombia). An. Inst. Invest. Mar Punta Betin 21: 117-125. [ Links ]
Taylor, W.R. & G.C. Van Dyke. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium, 9: 107-119. [ Links ]
Schultz, L.P. 1944. The fishes of the family characinidae from Venezuela, with description of seventeen new forms. Proc. U.S. Nat. Mus. 95: 235-367. [ Links ]
Vari, P.R. & D.J. Siebert. 1990. A new, unusually dimorphic species of Bryconamericus (Pisces:Ostariophysi:Characidae) from the Peruvian Amazon. Proc. Biol. Soc. Wash. 103: 516-524. [ Links ]
Weitzman, S.H. & S.V. Fink. 1985. Xenurobrycon phylogeny and putative pheromone pumps in glandulocaudine fishes (Teleostei: Characidae). Smith. Cont. Zool. (421): 1-118. [ Links ]
1 Universidad del Quindio, Departamento de Biología, A.A. 460, Armenia, Quindio, Colombia. Fax: (57) 7462563. Correo electrónico: croman@cocora.uniquindio.edu.co

