










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.48 n.1 San José Mar. 2000
a lluvias en la costa pacífica de Colombia
Andrés Velasco & Matthias Wolff 1
Recibido 11-I-1999. Corregido 16-IX-1999. Aceptado 22-IX-1999.
Abstract
Based on ichthyoplancton research along the southern Pacific coast of Colombia, which revealed high concentrations of fish larvae in this area, a juvenil fish survey was conducted in shallow waters (<10 meter water depth) during the transition period from dry to wet season (July 21 to July 31, 1994). The twelve sampling stations, were distributed in front of the four main of this coastline, where the highest concentrations of juvenile fish were expected. At each sample station, three 15 min oblique hauls were conducted every 8 hours using a bottom trawl for juvenile fish of 4 mm mesh size; 53 species in 20 families were identified (21 not known as adults in the area). They were distributed as follows: Sciaenids (11), Ariids (7), Pristigasterids (6), Engraulids (6), Carangids (5), Clupeids (2), Soleids (2), Batrachoidids (2), Tetraodontids (2), Urolophids (1), Polynemids (1), Synodontids (1) and Taeniopedids (1). This ichthyofauna is common in open tropical estuaries. Abundances and biomass were 0.35 ind./m3 and 1.61 g/m3 respectively, higher than in other tropical shallow water systems. The niches of the 25 species whose abundance (A) and biomass (B) contributed 97.5% and 93.3% respectively to the total catches were: (1) pelagic-neritic (53% A, 34% B); (2) demersal-pelagic (18% A, 28% B); (3) demersal (22% A, 23% B) and (4) benthic (8% A, 15% B). The Shannon-Weaver diversity index in the four subareas ranged from 0.89 to 1.1, within the range reported for other tropical American estuarine systems. An ordination analysis (non-metric Multi-Dimensional Scaling, MDS) of the data revealed significant differences in the species composition between day and night and an important association among some species. Station position, tides and environmental parameters did not markedly influence species composition. It is concluded that: (1) The area surveyed represents an important nursery and growth habitat for juvenile fish of many species that are more common as adults over soft bottoms in deep waters of the continental shelf; (2) this shallow soft bottom fish community is clearly separated from those species inhabiting intertidal salt marshes, interior bays and mangrove areas; and (3) some of the fish species commonly living in rocky and reef habitats in adult stages, spend their juvenile stage over shallow water soft bottoms. These results confirm a rather strong niche separation between juvenile and adults fish species stages in tropical waters.
Key words
Nurseries, juvenil fish assemblages, fish diversity, estuaries, multivariate analysis, Colombian Pacific Coast.
Desde finales de los años 80 se han realizado prospecciones dirigidas a evaluar los recursos pesqueros de la plataforma continental (20-200 m) de la costa pacífica colombiana en el marco de la cooperación técnica internacional (Anónimo 1987, 1988). El estudio de las aguas someras de menos de 20 m de profundidad, las cuales no fueron incluídas en las primeras expediciones, han comenzado a ser estudiadas más recientemente en el marco de la cooperación técnica europea para la pesca (Anónimo 1991, 1995).
Los estudios de López & Bussing (1982), Bartels et al. (1984), Bussing & López (1993, 1996) y Wolff (1996) constituyen los antecedentes recientes más importantes en el estudio de la ictiofauna adulta de la región del pacífico Centro-oriental. Un primer inventario de la ictiofauna marina adulta comercial a lo largo de la costa pacífica colombiana (Rubio 1988) arrojó 284 especies de 55 familias. El presente estudio tiene como objetivos: 1. Iniciar un inventario de las especies de juveniles de peces en las aguas someras (<10 m) frente a la parte sur (4°30N-77°21W y 1°53N-78°35W) de la costa pacífica colombiana, 2. Analizar posibles diferencias en la ocurrencia de ciertas especies de la comunidad de juveniles de peces a lo largo del gradiente geográfico y entre el día y la noche y 3. Comparar el espectro de especies de juveniles de peces capturados con el de la ictiofauna juvenil y adulta de fondos blandos registrada en la literatura.
A través de la comparación entre la composición de la fauna capturada de especies de juveniles de peces con la de la ictiofauna adulta de fondos blandos registrada en la zona, se pone a prueba la hipótesis de Yáñez-Arancibia & Sánchez-Gil (1988), la cual asume una mayor separación de los hábitats de juveniles y adultos en los ecosistemas tropicales que en los de latitudes más elevadas.
Materiales y Métodos
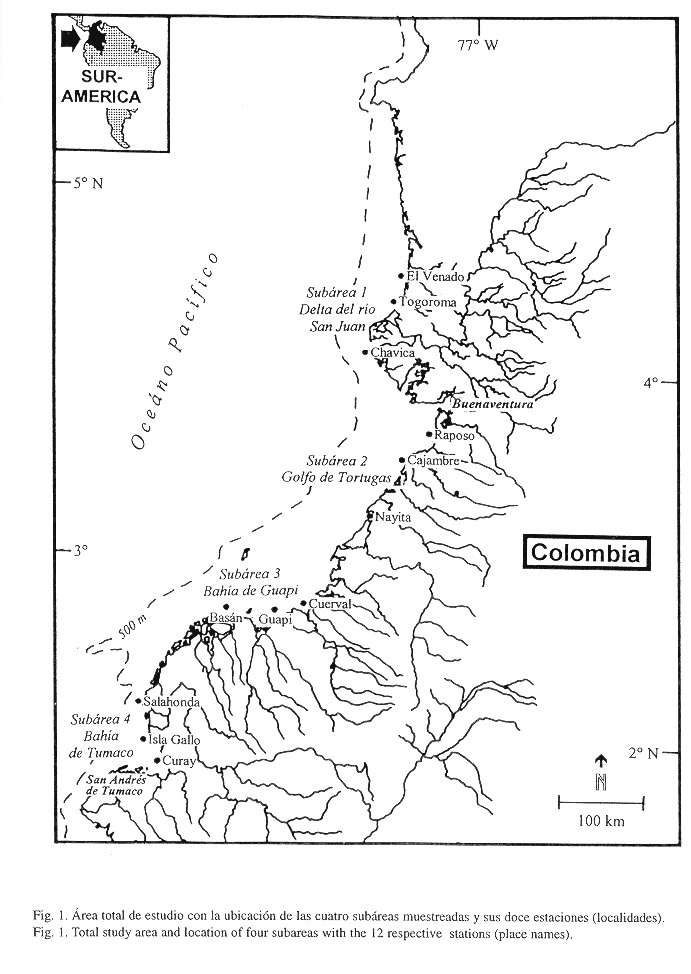
Área de estudio: El área de estudio se extiende desde los 04°3003N-77°2148W hasta los 01°5300N-78°3500W (Fig. 1) y comprende cuatro subáreas de: 1) Delta del río San Juan (estaciones El Venado, Togoroma y Chavica), 2) Golfo de Tortugas (estaciones Raposo, Cajambre y Nayita), 3) Bahía de Guapi (estaciones Cuerval, Guapi y Basán) y 4) Bahía de Tumaco (estaciones Salahonda, Isla Gallo y Curay). Las tres estaciones dentro de cada subárea están separadas cerca de 10 millas entre sí y se encuentran en aguas someras alrededor de una milla de la costa. El criterio para la selección de las 12 estaciones ubicadas en aguas someras frente a estuarios abiertos bordeados de manglares en la parte sur de esta costa fue el haber encontrado en ellas con anterioridad mayores concentraciones de ictioplancton (Rueda et al. 1992, Beltrán et al. 1994, Zapata et al. 1995).
Los 1300 km de costa pacífica colombiana pertenecen al Golfo de Panamá y sus partes norte y sur se diferencian topográfica, oceanográfica, climática e hidrográficamente. La parte sur del Pacífico colombiano se caracteriza por la presencia de 22 ríos (Ramírez & Bustos, 1976) que transportan cerca de 350 x 109 m3/año de sedimentos al área costera, los cuales junto con el clima cálido (promedio anual de 25-27°C, Sánchez et al. 1997) y una alta lluviosidad de 1 000-4 000 mm de precipitación al año favorecen el crecimiento de bosques de manglar (Prahl et al. 1990) altamente productivos (10-14 t/ha/año, Alvarez-León 1993).
Estrategia de muestreo y proceso de las muestras: La toma de muestras se efectuó del 21 al 31 de julio de 1994. Para ello se empleó un cúter del Instituto de Pesca y Acuicultura INPA (1.2 m de calado). Para las capturas se utilizó una red de investigación para juveniles de peces de 12 m de largo, 4.6 m2 de abertura de boca y 4 mm de ojo de malla en el copo, la cual fué arrastrada circularmente a media agua con ayuda de un cabrestante en aguas someras (<10 m de profundidad) sobre estrato arenoso-fangoso durante 10-15 min a una velocidad de cerca de 1.5 nudos. El volumen del agua filtrada se midió con un contador de flujo fijado a uno de los cabos de la red. En cada estación fueron realizados tres arrastres oblicuos con cerca de ocho horas de diferencia y se midieron los parámetros ambientales de temperatura, salinidad, oxígeno disuelto y profundidad.
Los peces se preseleccionaron por familias y/o especies y se fijaron en una solución de agua de mar al 7% de formol neutralizado con bórax. La identificación taxonómica de los peces se realizó en el laboratorio bajo un binocular con la ayuda de claves (Fischer 1978, Whitehead 1985, Whitehead et al. 1988; Rubio 1988), una lista (López & Bussing 1982) y una guía ilustrada de peces del Pacífico centroamericano (Bussing & López 1993, 1996).
Análisis de datos: Los valores de abundancia y biomasa de cada muestra fueron convertidos a 1000 m3 de agua filtrada. Las muestras tomadas entre las 06:00 y 18:00 hr fueron consideradas como diurnas y las tomadas entre las 18:00 y 06:00 hr como nocturnas.
La representatividad de las capturas de la comunidad de peces muestreada fué averiguada por medio de una curva del número de muestras contra el número acumulado de familias. La comparación de las cuatro subáreas muestreadas y la caracterización de la estructura de la comunidad de juveniles de peces en toda el área de muestreo, fueron efectuadas con base en los datos de abundancias y biomasas de las diferentes especies por medio de un análisis multivariado de las capturas. De este modo se pudieron comprobar diferencias día-noche en la composición específica de las capturas, como también describir determinadas asociaciones de especies. Posteriormente se compararon los espectros de la ictiofauna juvenil marino-estuarina encontrada con el de la ictiofauna adulta de fondos blandos registrada para la plataforma continental sobre estrato arenoso-fangoso.
Para el análisis multivariado se elaboraron sendas matrices de especies y estaciones basadas en el valor descriptivo de p = (B/A)0.73 x A, una expresión de la combinación de las abundancias y biomasas de Warwick & Clarke (1993), la cual relativiza la importancia de especies abundantes con bajas biomasas y especies raras con mayores pesos. Con la ayuda del programa PRIMER (Plymouth Routines In Multivariate Ecological Research) se calcularon las similaridades con base en el empleo del coeficiente de Bray-Curtis y se emplearon los métodos gráficos del análisis de dendrogramas y de escalamiento multidimensional no métrico NMDS (Field et al. 1982). El dendrograma de especies se elaboró con base en los valores de "p" de las 25 especies cuyas abundancias y biomasas representaron respectivamente 97.5% y 93.3% del total de las abundancias y biomasas. Para el dendrograma de estaciones se emplearon los valores de "p" de todas las pescas a nivel de familia, a fin de poder tener en cuenta las especies raras de grandes depredadores como congrios, barbetas, lenguados, peces lagarto, tembladores y rayas de aguijón. Los dendrogramas se elaboraron con valores de similaridad clasificados por rango para facilitar su comparación con los resultados de los gráficos NMDS (Clarke 1993). Por recomendación de Clarke & Warwick (1994a) se estandarizaron los datos debido a las marcadas diferencias de las abundancias y biomasas entre las muestras, y para evitar una sobreestimación del significado de las especies raras se transformaron los datos por medio de la aplicación de la raíz cuarta (Clarke & Green 1988). La determinación de diferencias significativas de las agrupaciones en los dendrogramas según grupos de especies, grupos de estaciones del día o la noche, o de marea baja o alta se efectuó por medio del test ANOSIM (Analysis of Similarities, Clarke & Warwick 1994b). Por medio del test SIMPER (Similarity Percentages, Clarke & Warwick 1994a) se averiguaron las familias de peces responsables de las agrupaciones día-noche en el dendrograma de estaciones. Los datos de los parámetros medioambientales de cada estación fueron correlacionados con los de las biomasas por medio del coeficiente por rangos de Spearmann. La diversidad se calculó de acuerdo con el índice numérico de Shannon & Weaver (1949) y se comparó con los valores registrados de otros estuarios americanos.
Resultados
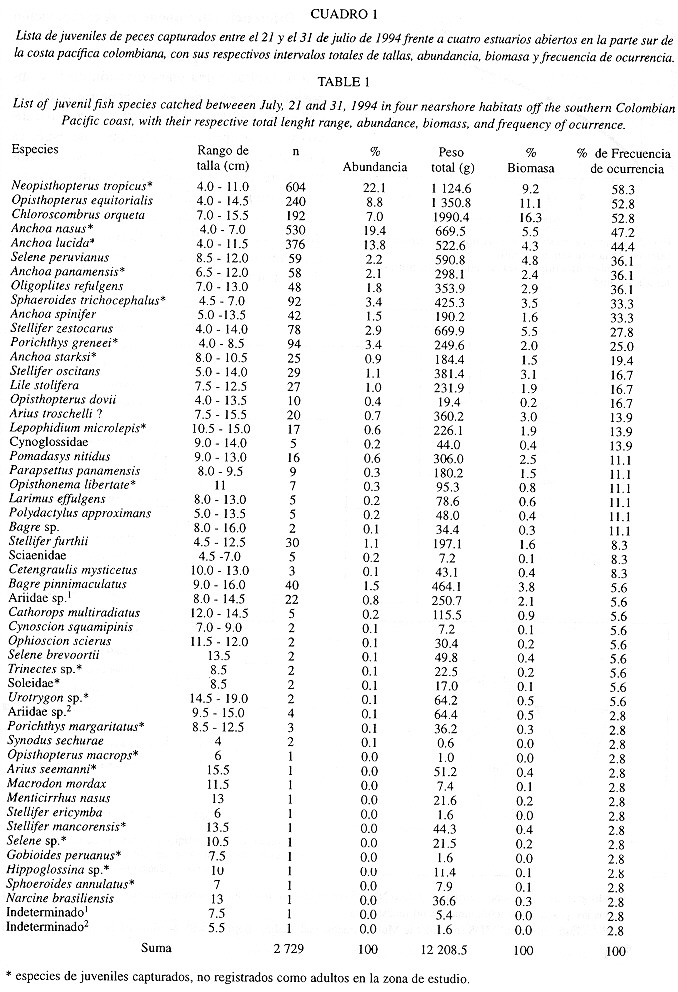
En total fueron identificados 2 729 juveniles de peces (intervalo total de tallas 4 a 16 cm) de 53 especies pertenecientes a 20 familias (Cuadro 1). Veintiún especies de trece familias no fueron hasta ahora registradas como adultos en el área de estudio. Las abundancias y biomasas promedio para toda el área de muestreo fueron de 0.35 Ind./m3 y 1.61 g/m3. Veiticinco especies de once familias sumaron 97.5% de las abundancias y 93.3% de las biomasas totales. Once especies estuvieron presentes en el 28% de todas las pescas. La relación entre el número de muestras y el número acumulado de familias indica que la ictiofauna juvenil capturada está bien representada en las 36 muestras (Fig. 2).
Diferencias día-noche en la composición de las capturas y asociaciones de especies: El análisis de dendrogramas y el NMDS dieron como resultado una clara división en la composición por especies de las capturas del día y de la noche (ANOSIM-Test: 0.4%, significativo) y tres grupos de asociaciones de especies (ANOSIM-Test: 0.0%, altamente significativo) pertenecientes a familias de diferentes nichos ecológicos (Fig. 3).
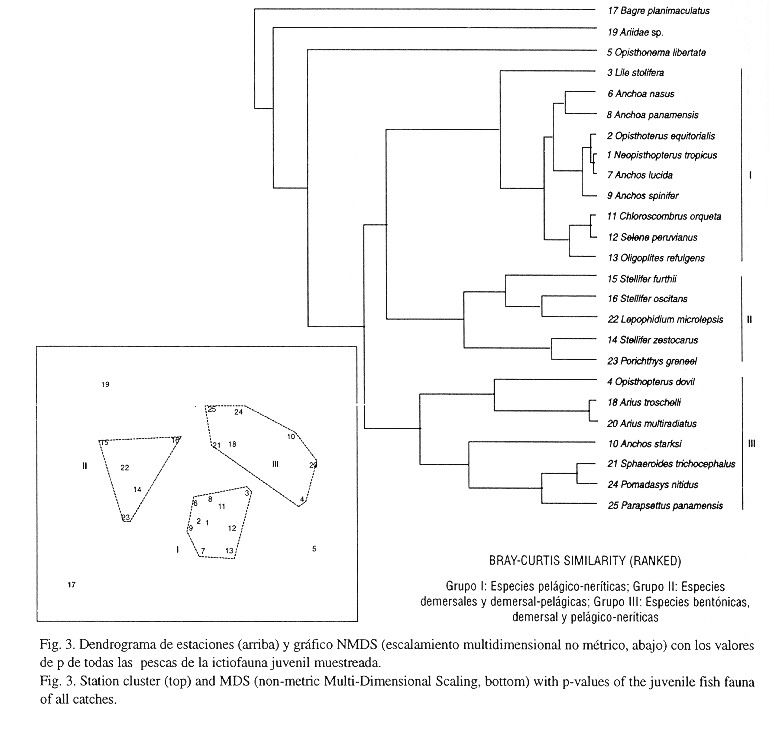
Las similaridades averiguadas por medio del test SIMPER arrojaron básicamente especies pelágico-neríticas en el día (grupo de estaciones en el centro del dendrograma) y pelágico-neríticas, demersal-pelágicas, demersales y bentónicas en la noche (grupos de estaciones superior e inferior) (Fig. 4). Las siguientes familias fueron responsables por la similaridad dentro de los tres grupos de estaciones (entre paréntesis la similaridad media en porcentaje): a) Grupo de estaciones nocturno de abajo (54.5%): Sciaenidae (26.8%), Carangidae (15.2%), Pristigasteridae (15.1%), Engraulidae (12.2%), Ariidae (12.9%) Tetraodontidae (7.63%) y Haemulidae (2.73%); b) Grupo de estaciones nocturno de arriba (46%): Pristigasteridae (46.8%), Engraulidae (37%) y Carangidae (1.9%), Batrachoididae (7.4%) y Sciaenidae (5.7%); c) Grupo de estaciones diurnas en el centro (57.2%): Carangidae (42.1%), Pristigasteridae (27.4%) y Engraulidae (27.3%).
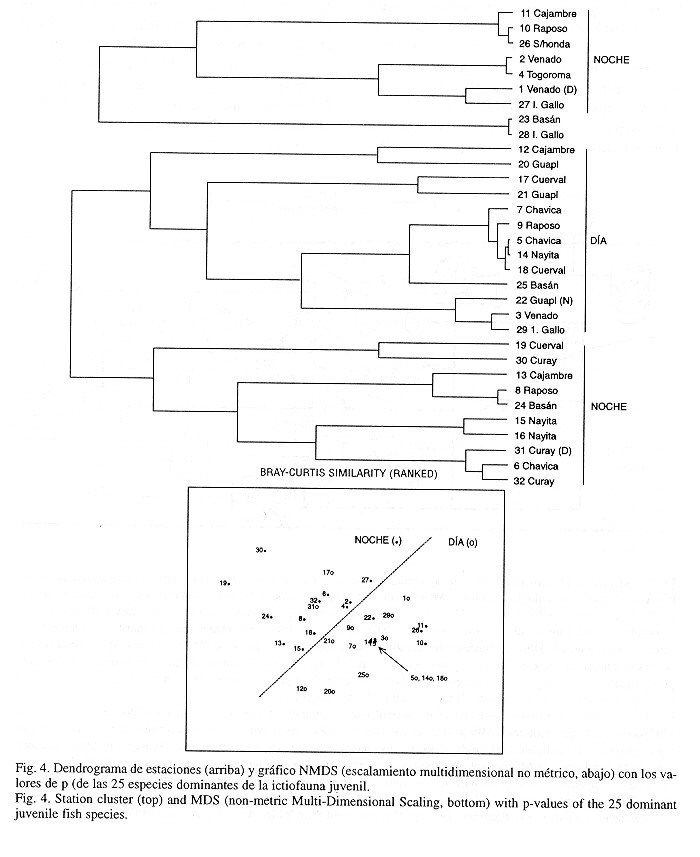
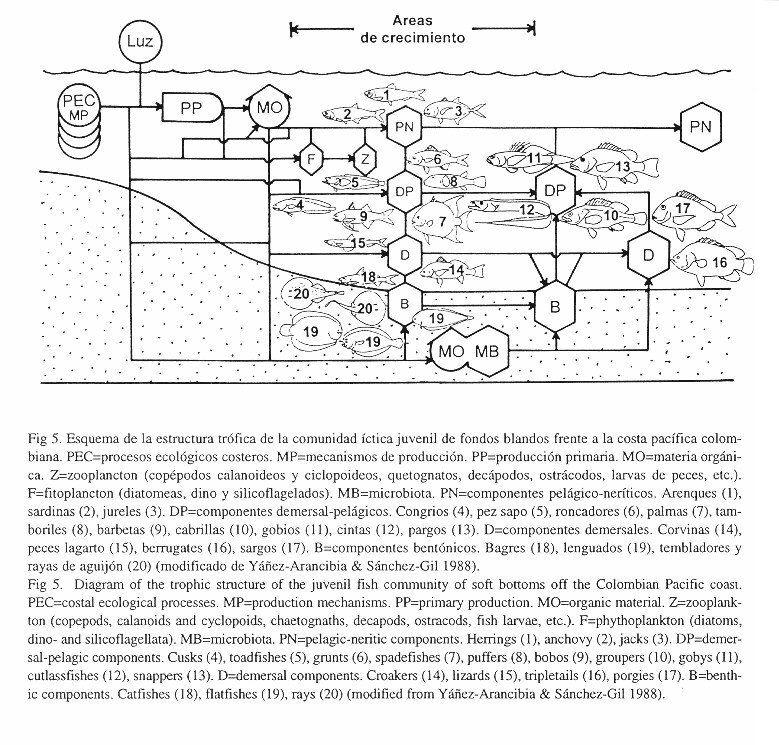
Las sardinas Opisthopterus equitorialis y Neopisthopterus tropicus formaron junto con las anchoas Anchoa lucida y A. spinifer y los jureles Chloroscombrus orqueta, Selene peruvianus y Oligoplites refulgens un primer grupo de especies pelágico-neríticas. Las corvinas Stellifer furthii, S. oscitans y S. zestocarus, formaron junto con el congrio Lepophidium microlepis y el pez sapo Porichthys greenei un segundo grupo de especies típicas de profundidad media (demersal-pelágicos). Los bagres (bentónicos) Arius troschelli y C. multiradiatus, el pez globo Sphoeroides trichocephalus, el roncador Pomadasys nitidus y la palma Parapsettus panamensis (demersal-pelágicos) formaron junto con la sardina Opisthopterus dovii y la anchoa Anchoa starksi (pelágico-neríticos) un tercer grupo de especies de toda la columna de agua (Fig. 3). Con base en todo lo anterior se ha bosquejado la estructura trófica de la comunidad de juveniles peces de fondos blandos en el sur de la costa pacífica colombiana (Fig. 5).
Discusión
El espectro de especies encontrado en los cuatro sistemas estudiados corresponde a una ictiofauna juvenil característica de aguas someras marino-estuarinas. Ella está conformada predominantemente por especies pelágico-costeras y demersales como ha sido descrita por Pauly (1985) para estuarios tropicales abiertos de fondos blandos. La mayor parte de las especies de ésta ictiofauna juvenil se encuentra como adulta, con algunas excepciones, sobre fondos arenoso-fangosos en aguas más profundas de la plataforma. Tal es el caso de diferentes especies de juveniles pertenecientes a las familias de las corvinas, bagres y rayas de aguijón, como jureles, tamboriles, peces perro y peces lagarto encontradas en las aguas del área de estudio entre 0-10 m de profundidad, las cuales se encuentran como adultos en el pacífico Centro-oriental a diferentes profundidades hasta los 100 m. Otras especies de juveniles encontradas en el presente trabajo migran como adultos aún a profundidades mayores de 100 m, como p.ej. el pez perro (Wolff 1996), lo cual sugiere que en el área de estudio se desarrollan también peces de hábitats más profundos. Algunas especies de juveniles encontradas viven como adultas sobre fondos rocosos o arrecifes de coral, como p.ej. el roncador Pomadasys nitidus, la palma, Parapsettus panamensis y el congrio Lepophidium microlepsis (Rubio 1988), u otros manglares, como p.ej. el tamboril Sphoeroides annulatus y el bagre Arius semanni (Szelistowski 1990).
De acuerdo con las diferencias encontradas en la composición de las capturas del día a la noche se asume una migración vertical nocturna de los juveniles de especies demersal-pelágicas (Sciaenidae, Tetraodontidae y Haemulidae), demersales (Batrachoididae) y béntónicas (Ariidae) en las aguas someras. La no significancia encontrada para las diferencias en la presencia de especies a lo largo del gradiente geográfico y en la correlación entre los datos de los parámetros medioambientales y los de las biomasas, se debe muy probablemente a las similitudes en la composición por especies frente a los cuatro estuarios abiertos estudiados y la homogeneidad de sus características medioambientales en el corto tiempo de la colecta.
La presencia en las capturas de juveniles de especies no registradas como adultos en la zona, como la ausencia de juveniles de especies eurihalinas de fondos blandos de los hábitats aledaños (charcos intermareales, manglares y bahías internas) registradas en la literatura, cuyos adultos prefieren otros tipos de sustrato, demuestran una clara separación de los hábitats de juveniles y adultos en el área de estudio confirmando la hipótesis de la separación de los habitats de los juveniles y los adultos (Fig. 5):
- especies demersales de la familia Lobotidae, como p.ej. berrugates (como juvenil en bahías internas o manglares y como adultos en áreas marino-estuarinas de fondo rocoso o arenoso);
- Xenichthyidae (Palometas): como juveniles en charcos intermareales de fondo rocoso y como adultos en arrecifes de coral (Rubio 1988);
- peces de aguas profundas de la plataforma como p.ej. cabrillas (Serranidae): como juveniles en bahías internas y estuarios y como adultos sobre fondo rocosos y arrecifes; gobios (Gobiidae) y cintas (Trichiuridae): como juveniles en los manglares (Yáñez-Arancibia et al. 1985 y Yáñez-Arancibia & Sánchez-Gil 1986);
- peces del manglar como p.ej. pargos (Lutjanidae): como juveniles en charcos intermareales; Cyprinodontidae y Gobiosocidae (Szelistowski 1990);
- peces de lagunas costeras como p.ej. lisas (Mugilidae): como juveniles de más de 3 cm incluso en aguas dulces de ríos y arroyos (Velasco & Villegas 1985); Gerreidae (Mojarras): como juveniles en aguas dulces; róbalos (Centropomidae): juveniles en la época de lluvias en charcos intermareales (Rubio 1988).
Estructura de la comunidad:Si bien las especies de juveniles capturadas reflejan todo el espectro adulto a nivel de familia reportado para las aguas someras de estrato arenoso-fangoso en el área de estudio, es de suponer que los predadores, en su mayoría grandes demersales y especies bentónicas, estén subestimados en las capturas con los arrastres a media agua. Ello explica en parte que los congrios (Lepophidium microlepsis), barbetas (Polydactylus approximans), lenguados (Cynoglossidae, Trinectes sp. e Hipoglossina sp.), peces lagarto (Synodus sechurae), tembladores (Narcine brasiliensis) y rayas de aguijón (Urotrygon sp.) estuvieron representados apenas en un 2.5% de las abundancias y 6.7% de las biomasas totales. Los consumidores secundarios y terciarios, como los bagres (Bagre pinnimaculatus, Ariidae spp., Arius troschellli, C. multiradiatus y A. seemanni) y corvinas (Stellifer zestocarus, S. furthii, S. oscitans und S. mancorensis) estuvieron sin embargo mayormente representados respectivamente en las estaciones con los mayores aportes fluviales de los ríos Patía y Mira (las estaciones Cuerval y Basán en la Bahía de Guapi y las estaciones Curay e Isla Gallo en la Bahía de Tumaco). No obstante, un estimativo de la biomasa media de juveniles en la columna de agua del área de estudio de aproximadamente 8 g/m2 es relativamente alto comparado con otros valores de biomasa en aguas marino-estuarinas tropicales como p.ej. de Longhurst (1957) para aguas costeras de Sierra Leona (7 g/m2), de Mines et al. (1986) para la bahía de San Miguel en las Filipinas (2.13 g/m2), y de Lohmeier (1987) para aguas someras tropicales en Indonesia (6 g/m2).
CUADRO 2
Indices de diversidad (Hs) de algunas comunidades ictiofaunísticas de estuarios americanos.
TABLE 2
Diversity index (Hs) of several communities from different American estuaries.
| | | | | |
| | | | | |
| California central | | | | |
| Sur de California | | | | |
| Puerto de Los Angeles | | | | |
| Bahía de Galveston | | | | |
| Estuario del río Mystic. USA | | | | |
| Golfo de Nicoya | | | | |
| Golfo de Nicoya | | | | |
| Golfo de Nicoya | | | | |
| Pacífico mejicano | | | | |
| Laguna de Términos | | | | |
| Bahía de Jiquilisco | | | | |
| Bahía de Panamá | | | | |
| San Juan Delta | | | | |
| Golfo de Tortugas | | | | |
| Bahía de Guapi | | | | |
| Bahía de Tumaco | | | | |
| Pacífico SO colombiano | | | | |
Claras variaciones en los parámetros medioambientales descritas para el área en la literatura para la época de transición de la época de lluvias a la época seca en el mes de julio, como lo son el ascenso de los valores de salinidad, temperatura y transparencia del agua, el descenso de la materia orgánica disuelta transportada por los numerosos ríos (POM) (Martínez & Carvajal 1990) y el debilitamiento de la Corriente Colombia (Wooster 1959) marcan muy probablemente un reclutamiento de un mayor número de especies de peces en la época de transición de lluvias a secas que en el resto del año en el área de estudio, lo que se ha llamado hasta ahora empíricamente época de juveniles.
Agradecimientos
Los resultados de este estudio forman parte de una tesis doctoral de la Universidad de Bremen, financiada por el Servicio Alemán de Intercambio Académico (DAAD). Los autores agradecen a Eduardo Del Real del Programa de Pesca UE/VECEP ALA 92/43 y Juan Valverde del Instituto Colombiano de Pesca y Acuicultura (INPA) por el apoyo logístico y financiero para el muestreo en Colombia.
Resumen
En base a mayores concentraciones de plancton encontradas en la parte sur de la costa pacífica colombiana, se colectó su ictiofauna juvenil en aguas someras (<10 m de profundidad) en la época de transición de secas a lluvias, frente a sus cuatro estuarios más importantes. El área de estudio comprendió 12 estaciones. En cada estación, se efectuaron entre el 21 y el 31 de julio de 1994 tres arrastres oblicuos de 15 min de duración, con aprox. ocho horas de diferencia, utilizando una red para juveniles de peces de 4 mm de ojo de malla en el copo. En total fueron capturadas 53 especies pertenecientes a 20 familias (21 de ellas no registradas hasta ahora como adultos en la zona). Estas se distribuyeron como sigue: Sciaenidae (11), Ariidae (7), Pristigasteridae (6), Engraulidae (6), Carangidae (5), Clupeidae, Soleidae, Batrachoididae, Tetraodontidae (2 especies cada una), Bothidae, Cynoglossidae, Ophidiidae, Haemulidae, Ephipidiidae, Torpedinidae, Urolophidae, Polynemidae, Synodontidae y Taeniopedidae (1 especie). Esta ictiofauna es característica de estuarios abiertos. Las abundancias y biomasas medias fueron 0.35 Ind./m3 y 1.61 g/m3 respectivamente, valores comparativamente altos para aguas costeras tropicales. El agrupamiento de las 25 especies cuyas abundancias (A) y biomasas (B) representaron respectivamente 97.5% y 93.3% del total de las capturas, de acuerdo a sus nichos ecológicos, arrojó la siguiente composición: 1. Pelágico-neríticas (53% A; 34% B) 2. Demersal-pelágicas (18% A; 28% B) 3. Demersales (22% A; 23% B) y 4. Bentónicas (8% A; 15% B). El índice de diversidad de Shannon-Weaver en las cuatro subáreas, varió entre 0.89 y 1.1 encontrándose dentro del rango reportado para otros estuarios americanos. La ordenación de los datos de las capturas (escalamiento multidimensional no métrico, NMDS) reveló diferencias día/noche en la composición específica y determinadas asociaciones de especies. La ubicación geográfica de las estaciones, la marea y los parámetros ambientales no jugaron un papel significativo en su distribución. Se concluye que: 1) el área estudiada representa una zona de crecimiento de una ictiofauna adulta típica de fondos blandos de aguas más profundas de la plataforma continental; 2) esta comunidad está claramente separada de la comunidad de juveniles de peces eurihalinos de fondos blandos típicos de charcos intermareales, manglares y bahías interiores y 3) algunas especies cuyos estadíos juveniles transcurren sobre fondos blandos de las aguas someras, migran como adultas a hábitats rocosos o coralinos. Lo anterior confirma una mayor separación por nichos entre juveniles y adultos, para las especies de peces de aguas tropicales.
Referencias
Allen, G. & H. Horn. 1975. Abundance diversity and seasonality of fishes in Colorado Lagoon, Alamitos Bay, California. Estuarine Coast. Mar. Sci. 3: 185-242. [ Links ]
Alvarez-León, R. 1993. Mangrove ecosystems of Colombia: ITTO/ISME Technical Report Project PD 114/90(F), Part I Latin America. International Tropical Timber Organization and International Society for Mangrove Ecosystems, Yokohama, Japón. 38 p. [ Links ]
Araya, H. 1988. Diversidad, distribución, abundancia y relaciones tróficas de peces en Estero Damas y Estero Palo Seco. Aguirre (Quepos), Parrita, Puntarenas. Costa Rica. Tesis de Licenciatura en Biología, Universidad de Costa Rica, San José, Costa Rica. [ Links ]
Bartels, C.E., K.S. Price, M.L. Bussing & B.A. Bussing. 1984. Ecological assessment of finfish as indicators of habitats in the Gulf of Nicoya, Costa Rica. Hidrobiologia 112: 197-207 p. [ Links ]
Bechtel, T.J. & B.J. Copeland. 1970. Fish species diversity indices as indicators of pollution in Galveston Bay, Texas. Contrib. Mar. Sci. 15: 103-132. [ Links ]
Beltrán León, B.S., G.E. Ramos Tafur, J. C. Escobar Valderrama & J.A. Tovar. 1994. Distribución y abundancia de huevos y larvas de Opisthonema sp. (Gill, 1861) y Centegraulis mysticetus (Günther, 1866) (Pisces: Clupeiformes) en el Pacífico colombiano durante enero de 1993. Bol. Cient. INPA (2): 181-199 p. [ Links ]
Bussing, W. A. & M. I. López. 1993. Demersal and Pelagic Inshore Fishes of the Pacific Coast of Lower Central America. Rev. Biol. Trop., San José, Costa Rica. 163 p. [ Links ]
Bussing, W. A. & M. I. López. 1996. Fishes collected during the Victor Hensen Costa Rica Expedition (1993/1994). Spec. Pub. Rev. Biol. Trop. 44: 183-186 p. [ Links ]
Anónimo. 1991. Cruceros hidroacústicos de evaluación de peces pelágicos pequeños en el Pacífico colombiano. Programa regional de cooperación técnica para la pesca Comunidad Económica Europea/Perú, Ecuador y Colombia, Bogotá, Colombia. CEE/PEC.17 p. [ Links ]
Clarke, K.R. 1993. Non-parametric multivariate analysis of changes in community structure. Austral. J. Ecol. 18: 117-143. [ Links ]
Clarke, K.R. & R.H. Green. 1988. Statistical design and analysis for a biological effects study. Mar. Ecol. Prog. Ser. 46: 213-226. [ Links ]
Clarke, K.R. & R.M. Warwick. 1994a. Change in marine communities: an approach to statistical analysis and interpretation. Natural Environment Research Council, Plymouth, Inglaterra. 144 p. [ Links ]
Clarke, K.R. & R.M. Warwick. 1994b. Similarity-based testing for communinity pattern: the two-way layout with no replication. Mar. Biol. 118: 167-176. [ Links ]
DCroz, L. & A. Averza. 1979. Observaciones sobre la abundancia y diversidad de las poblaciones de peces estuarinos en el Caribe de Panamá. Rev. Biol. Trop. 27: 189-201. [ Links ]
Field, J.G., Clarke, K.R. & R.M. Warwick. 1982. A practical strategy for analysing multispecies distribution patterns. Mar. Ecol. Prog. Ser. 8: 37-52. [ Links ]
Fischer, W. 1978. FAO species identification sheets for fishery purposes. Western Central Atlantic (fishing area 31). FAO, Roma, 150 sheets. [ Links ]
Haedrich, R.L. & S.O. Haedrich. 1974. A seasonal survey of the fishes in the Mystic River, a polluted estuary in downtown Boston, Massachusetts. Estuarine Coastal Mar. Sci. 2: 59-73. [ Links ]
Hernández & P. Chavance. 1985. Ecología de poblaciones de peces dominantes en estuarios tropicales: Factores ambientales que regulan las estrategias biológicas y la producción, p. 311-366. In A. Yáñez-Arancibia (ed.). Fish community ecology in estuarine and coastal lagoons: Towards an ecosystem integration. Univ. Nal. Autón. Mexico. [ Links ]
Horn, M.H. 1980. Diel and seasonal variation in abundance and diversity of shallow - water fish populations in Morro Bay, California. Fish. Bull. 78: 759-770. [ Links ]
Anónimo. 1988. Informe de la investigación sobre recursos pesqueros marítimos en la República de Colombia. Informe Técnico Agencia de Cooperación Internacional del Japón, Tokio. 509 p. [ Links ]
León, P.E. 1973. Ecología de la ictiofauna del Golfo de Nicoya, Costa Rica, un estuario tropical. Rev. Biol. Trop. 21: 5-30. [ Links ]
Lohmeier, U.P. 1987. Bestandskundliche Untersuchungen der Bodenfische des östlichen Indischen Ozeans im vorgelagerten Schelf der Inseln Sumatra, Jawa, Bali. Dissertation, Universität Kiel, Kiel, Alemania. [ Links ]
Longhurst, A. 1957. Density of marine benthic communities off West Africa. Nature 179: 542-543. [ Links ]
López M. I. & W. A. Bussing. 1982. Lista provisional de los peces marinos de la Costa Pacífica de Costa Rica. Rev. Biol. Trop. 30: 5-26. [ Links ]
Martínez J.O. & J.H. Carvajal. 1990. Atlas de geomorfología y erosión de la costa pacífica colombiana (Valle, Cauca, Nariño). INGEOMINAS-PROGOG, Bogotá, Colombia. 167 p. [ Links ]
Mines, A.N., Smith, I.R. & D. Pauly. 1986. An overview of the fisheries of San Miguel Bay, Philippines, p. 385-388. In J.L.Mc Lean, L.B. Dizon und L.V. Hosillos (eds.). The First Asian Fisheries Forum. Asian Fisheries Society. Manila, Filipinas. [ Links ]
Anónimo. 1987. Reports on Surveys with R/V Dr. Fridtjof Nansen: Prospecciones de los recursos pesqueros de la plataforma pacífica entre el sur de México y Colombia.NORAD/UNDP/FAO. Inst. of Mar. Research. Bergen, Noruega. 96 p. [ Links ]
Pauly, D. 1985. Ecology of coastal and estuarine fishes in Southeast Asia: A Philippine case study, p. 499-514. In A. Yáñez-Arancibia (ed.). Fish community ecology in estuarine and coastal lagoons: Towards an ecosystem integration. Univ. Nal. Autón. México. México, D.F. [ Links ]
Phillips, P.C. 1981. Diversity and fish community structure in a Central America mangrove embayment. Rev. Biol. Trop. 29: 227-236. [ Links ]
Prahl, H. von, Cantera, J.R. & R. Contreras. 1990. Manglares y hombres del Pacífico colombiano. FEN-COLCIENCIAS, Bogotá, Colombia. 193 p. [ Links ]
Protti M. 1993. Dinámica estacional de la comunidad de peces en el interior del Golfo de Nicoya, Costa Rica. Tesis de Licenciatura en Biología, Universidad Nacional, Heredia, Costa Rica. [ Links ]
Ramírez, A. & N. Bustos. 1976. Estudio preliminar de precipitación y caudales medios de la vertiente del Pacífico. Tesis de grado, Universidad Jorge Tadeo Lozano, Bogotá, Colombia. [ Links ]
Rubio, E.A. 1988. Peces de importancia comercial para el Pacífico colombiano. Universidad del Valle, Cali, Colombia. 495 p. [ Links ]
Rueda, C., B. E. Beltrán, F. E. Delgado & A. Velasco. 1992. Ictioplancton de las Familias Clupeidae y Engraulididae en el Pacífico colombiano. Cruceros PEC-PELAG 1991. Programa regional de cooperación técnica para la pesca CEE/PEC/MINAGRICULTURA ALA 87/21. Buenaventura, Colombia. 123 p. [ Links ]
Sánchez Páez, H., Alvarez-León, R., Guevara Mancera, O.A., Zamora Guzmán, A., Rodríguez Cruz, H. & H.E. Bravo Pazmiño, 1997. Diagnóstico y zonificación preliminar de los manglares del Pacífico de Colombia. Informe técnico Proyecto PD/171/91 Rev. 2 (F) Fase 1, Organización Internacional de Maderas Tropicales, Ministerio del Ambiente y Asociación Colombiana de Reforestadores, Bogotá, Colombia. 343 p. [ Links ]
Shannon, C.E. & W. Weaver. 1949. The mathematical theory of communication. Univ. of Illinois Press, Urbana. 119 p. [ Links ]
Stephens, J.S.Jr., C. Terry, S. Subber & M.J. Allen. 1974. Abundance, distribution, seasonality and productivity of the fish populations in Los Angeles Harbor, 1972-73, p. 1-42. In Soule & Oguri (eds.). Marine Studies of San Pedro Bay, Part IV, Environmental Field Investigations, Allan Hancock Foundation Pub. USG-SG-6-72. [ Links ]
Szelistowski, W.A. 1990. Importance of mangrove plant litter in fish food webs and as temporary, floating habitat in the Gulf of Nicoya. Ph.D. thesis , University of Southern California. [ Links ]
Anónimo. 1995. Boletín VECEP Informa Nr. 1. Programa de pesca Unión Europea/Venezuela, Colombia, Ecuador y Perú, Lima, Perú. UE/VECEP 18 p. [ Links ]
Velasco Mora, A. & P.Z. Villegas. 1985. Aportes al conocimiento de la historia de vida de Mugil incilis (Hancock, 1830), Mugil liza (Valenciennes, 1836) y Mugil curema (Valenciennes, 1836) en la Ciénaga de Tesca. Tesis de grado, Universidad Jorge Tadeo Lozano, Bogotá, Colombia. [ Links ]
Warwick, R.M. & K.R. Clarke. 1993. Comparing the severity of disturbance: a metaanalysis of marine macrobenthic community data. Mar. Ecol. Prog. Ser. 92: 221-231. [ Links ]
Whitehead, P.J.P. 1985. Clupeoid Fishes of the World. FAO Fish. Synop. No. 125, Vol. 7, Part 1. Roma. 1-303 p. [ Links ]
Whitehead, P.J.P., G.J. Nelson and T. Wongratana. 1988. Clupeoid Fishes of the World. FAO Fish. Synop. No. 125, Vol. 7, Part 2. Roma. 305-579 p. [ Links ]
Wolff, M. 1996. Demersal fish assemblages along the Pacific coast of Costa Rica: a quatitative and multivariate assesment based on the Victor Hensen Costa Rica Expedition (1993/1994). Rev. Biol. Trop. Vol. 44 (18): 187-214. [ Links ]
Wooster, W.S. 1959. Oceanic observations in the Panama Bight, "Ascoy" expedition, 1941. Bull. Amer. Mus. Nat. Hist. 118: 113-152. [ Links ]
Yáñez-Arancibia, A. 1975. Sobre los estudios de peces en las lagunas costeras: Nota científica. Anal. Centro Cienc. del Mar y Limnol. Univ. Nal. Autón. México. 2: 53-60. [ Links ]
Yáñez-Arancibia, A., F. Amezcua-Linares & J.W. Day. 1980. Fish community structure and function in Términos Lagoon. A tropical estuary in the southern Gulf of México, p. 465-482. In V. Kennedy (ed.) Estuarine Perspectives. Academic, Nueva York. [ Links ]
Yáñez-Arancibia, A. 1984. Evaluación de la pesca demersal costera. Ciencia y Desarrollo CONACYT, 58 (X): 61-71. [ Links ]
Yáñez-Arancibia, A. 1985. Recursos demersales de alta diversidad en las costas tropicales: Perspectiva ecológica, p.17-38. In Yáñez-Arancibia, A. (ed.). Recursos pesqueros potenciales de México: La pesca acompañante del camarón. Prog. Univ. de Alimentos, Inst. Cienc. del Mar y Limnol., Inst. Nal. de Pesca. Univ. Nal. Autón. México. [ Links ]
Yáñez-Arancibia, A., A.L. Lara Domínguez, A. Aguirre León, S. Díaz Ruíz, F. Amezcua- Linares, D. Flores Hernández & P. Chavance. 1985. Ecología de poblaciones de peces dominantes en estuarios tropicales: Factores ambientales que regulan las estrategias biológicas y la producción, p. 311-366. In A. Yáñez-Arancibia (ed.). Fish community ecology in estuarine and coastal lagoons: Towards an ecosystem integration. Univ. Nal. Autón. México, México, D.F. [ Links ]
Yáñez-Arancibia, A. 1986. Ecología de la zona costera: Análisis de siete tópicos. AGT. México. 190 p. [ Links ]
Yáñez-Arancibia, A. & P. Sánchez-Gil. 1986. Los peces demersales de la plataforma sur del Golfo de México. Inst. Cienc. Mar Limnol. Univ. Nal. Autón. México, Publ. Esp. 9:1-230. [ Links ]
Yáñez-Arancibia, A. & P. Sánchez-Gil. 1988. Ecología de los recursos demersales marinos. AGT, México. 228 p. [ Links ]
Zapata Padilla L.A., B.S. Beltrán León & J.A.Tovar. 1995. Crucero de evaluación de huevos, larvas y adultos de pequeños pelágicos INPA/VECEP 9412. Programa regional de pesca INPA/VECEP ALA 92/43. Buenaventura, Colombia. 23 p. [ Links ]
1 Centro de Ecología Marina Tropical (ZMT), Fahrenheitstr. 1, 28359 Bremen, Alemania. Fax 04212208330. Internet: avelasco@uni-bremen.de mwolff@zmt.uni-bremen.de

