










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.48 n.1 San José Mar. 2000
Palinuridae) en el sureste del golfo de California, México
Isabel R. Muñoz-García Raúl Pérez-González Luis Miguel Flores-Campaña y Martín Ignacio Borrego 1
Recibido: 8-I-1998. Corregido: 19-VII-1999. Aceptado: 6-IX-1999.
Abstract
The spatial and temporal distribution of the stage I phyllosoma larvae of the lobsters Panulirus inflatus and P. gracilis and their relationships to water temperatures and salinities were analyzed from February to December 1989 in the southeastern Gulf of California, Mexico. Three stations were located within the bay of Mazatlan and one was located at the mouth of the Urias system (station one). Two-hundred and eight samples (104 at surface and 104 at bottom) were obtained from 26 horizontal plankton tows. Water temperatures and salinities were measured concurrently. Phyllosoma larvae were most abundant in station one, with total mean densities of 1 206 larvae/1 000 m3 (surface) and 1 196 larvae/1 000 m3 (bottom). Water temperatures ranged from 17.9°C, in February, to 30.2°C, in August, and water salinities ranged from 34.10/00, in August, to 35.00/00, in May. The abundance of phyllosoma larvae was not related to changes in water temperature or salinity, however it was significantly related with the reproductive behavior of adults inhabiting the area. Spatial distribution of phyllosoma strongly depends upon current patterns and tidal cycles.
Key words
Distribution, abundance, phyllosoma Panulirus, southeastern Gulf of California.
Las langostas espinosas Panulirus inflatus (Bouvier, 1895) y P. gracilis Streets, 1871 se distribuyen conjuntamente en las costas mexicanas del Pacífico (Holthuis 1991). Estas especies representan una pesquería artesanal importante en las costas del sureste del golfo de California. Sin embargo, las investigaciones realizadas en esta región son escasas y han sido dirigidas, principalmente, hacia la biología y la pesquería de las poblaciones de juveniles y adultos (Pérez-González et al. 1992a, 1992b, Flores-Campaña et al. 1993).
El ciclo biológico de las langostas espinosas es muy extenso e incluye una etapa larvaria denominada filosoma, la cual permanece derivando en el plancton de 6 a 11 meses (Johnson 1960, Buesa 1969, Chittleborough y Thomas 1969). Durante este período se presentan 11 estadíos larvales en P. interruptus (Randall, 1840) (Johnson 1956), P. inflatus (Johnson y Knight 1966) y P. gracilis (Báez 1983), antes de alcanzar la fase de puerulo. En las costas mexicanas del Pacífico, son pocos los estudios sobre la fase filosoma. Estos han sido realizados en aguas costeras y oceánicas (de 37 a 370 km de la costa y entre 50 y 2 500 m de profundidad) y versan sobre las características morfológicas, dispersión y abundancia de las filosomas (Johnson 1956, 1971, Johnson y Knight 1966). Sin embargo, en algunas especies como P. cygnus George 1962, Jasus edwardsii (Hutton,1875), P. marginatus (Quoy y Gaimard, 1825), Homarus americanus H. Milne Edwards, 1837 y P. argus (Latreille, 1804) se han realizado varias investigaciones sobre esta fase y en ellas se ha relacionado su dispersión y abundancia con la temperatura, salinidad, luz, hembras desovantes y con los patrones de corrientes litorales y de mareas de las zonas analizadas (Phillips 1981, Pearce y Phillips, 1988, Katz et al. 1994, Kittaka 1994, Rothlisberg et al. 1994, Alfonso et al. 1995).
En el caso de los recursos pesqueros, especialmente aquéllos que presentan un elevado valor en el mercado, como es el caso de las langostas, es relevante conocer los aspectos biológicos y ecológicos de todas las fases del ciclo de vida. Este conocimiento permite proponer su administración adecuada y racional. Debido a esto y a la ausencia total de estudios sobre la etapa filosoma de P. inflatus y P. gracilis en el sureste del golfo de California, el presente estudio tiene como finalidad evaluar su distribución y abundancia y su relación con la temperatura y la salinidad en aguas someras (6-19 m de profundidad), así como analizar su dispersión con respecto al patrón de las corrientes litorales y de mareas. Asimismo, se discute la relación entre los porcentajes de hembras ovígeras obtenidos por Pérez-González et al. (1992b) en la misma área y la presencia y abundancia de filosomas.
Material y Métodos
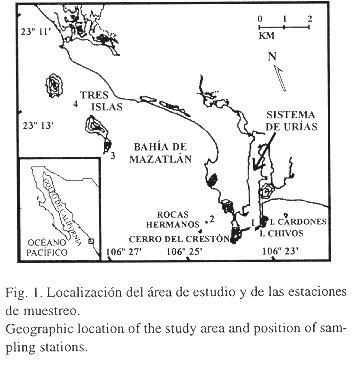
Entre febrero y diciembre de 1989 se realizaron 26 muestreos de plancton en superficie y fondo, en tres estaciones ubicadas en la bahía de Mazatlán (estaciones dos, tres y cuatro) y una en la boca del sistema de Urías (estación uno). Esta zona está localizada al sur del estado de Sinaloa entre los 23°08'48" y 23°16'00" N y los 106°29'24" y 106°23'26" W (Fig. 1) y es parte de una masa de agua costera influenciada por el régimen oceanográfico del sur del golfo de California, con una ligera estratificación termo-halina (Anónimo 1980).
Los muestreos se realizaron entre las 07:00 y las 11:00 hr e incluyeron arrastres horizontales de plancton de 5 min, a una velocidad aproximada de 2 nudos, con una red cónica de 2.20 m de largo, 0.58 m de diámetro de la boca y 475 mm de luz de malla. En la boca de la red se colocó un medidor de flujo previamente calibrado (1 revolución= 0.068 m3 de agua filtrada). El volumen de agua filtrado en cada arrastre varió entre 21.76 y 193.80 m3.
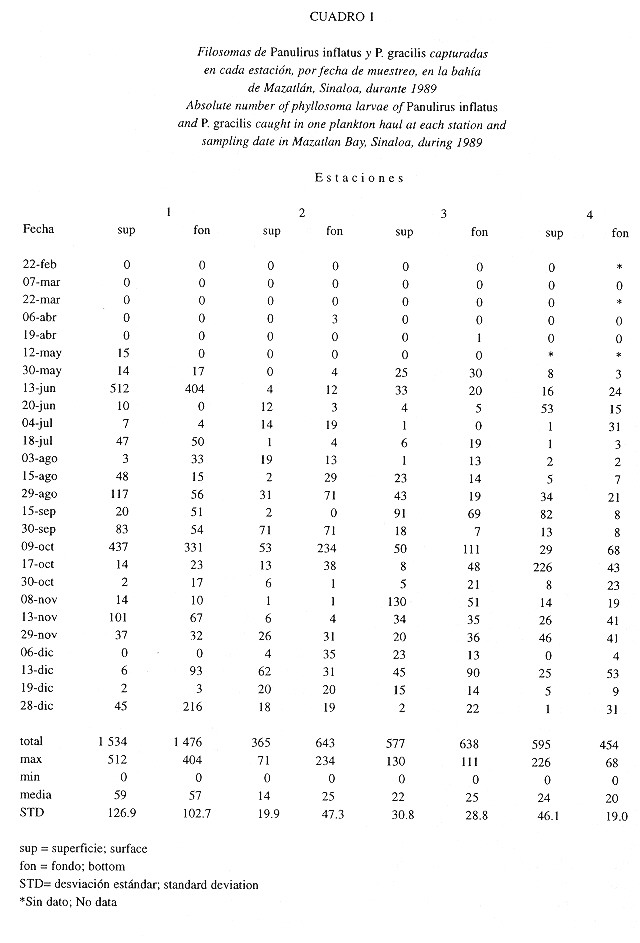
Se obtuvieron 208 muestras (104 de superficie y 104 de fondo). Los organismos recolectados se fijaron en formol al 5%. Las muestras se revisaron con un microscopio estereoscópico y las filosomas se separaron de acuerdo a las características descritas por Johnson y Knight (1966).
Las temperaturas de superficie y fondo se determinaron con un termómetro de cubeta de -1 a 51°C (± 0.1°C). La toma de las muestras de agua para medir la temperatura y la salinidad del fondo se realizó por medio de una botella Van Dorn. La determinación de la salinidad se hizo en el laboratorio con un salinómetro de inducción (± 0.0040/00) y la profundidad se estimó con una sondaleza manual.
Para establecer las diferencias entre la densidad de filosomas a través del período de estudio y entre las distintas zonas de muestreo, se efectuó un análisis de varianza no paramétrico de dos vías. En aquellos casos en que este análisis resultó significativo, se aplicó el método por rangos de Conover (1980) para determinar las diferentes agrupaciones.
Se aplicó un análisis de correlación para determinar la relación entre la densidad de larvas con la temperatura y la salinidad. Con los datos obtenidos por Pérez-González et al. (1992b), se realizó una comparación gráfica del porcentaje de hembras ovígeras con la densidad media mensual de filosomas.
Resultados
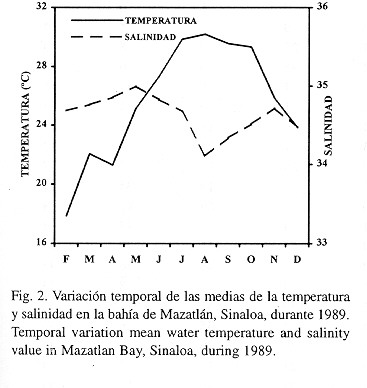
Los valores de la temperatura mostraron que en esta zona se presentan dos períodos: una época fría de diciembre a mayo (17.9-25.2°C) y una cálida de junio a noviembre (25.9-30.2°C) (Fig. 2). Las oscilaciones de la salinidad fueron menos marcadas que las de la temperatura, fluctuando de 34.11 en agosto (época de lluvias) a 35.00 0/00 en mayo (época de secas) (Fig. 2).
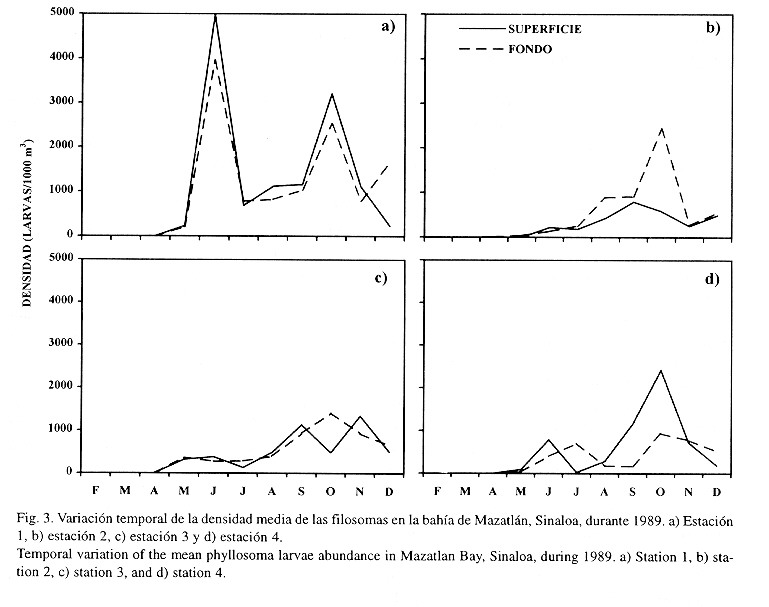
Se recolectaron un total de 6 282 filosomas, todas en el estadío I (Cuadro 1) y su presencia se detectó a partir de abril (con un valor máximo de 32 larvas/1 000 m3) hasta diciembre (con una densidad máxima de 4 963 larvas/1 000 m3) (Fig. 3). Las mayores densidades fueron obtenidas en la estación uno y se presentaron dos pulsos en la superficie: uno en junio (5 001 larvas/1 000 m3) y el otro en octubre (3 128 larvas/1 000 m3). En el fondo fueron observados tres pulsos (4 069 en junio, 2 742 en octubre y 1 669 larvas/1 000 m3 en diciembre) (Fig. 3a). En la estación dos las densidades máximas en la superficie y en el fondo fueron de 813 (en septiembre) y 2 607 larvas/1 000 m3 (en octubre), respectivamente (Fig. 3b). En la estación tres, las más altas densidades en la superficie se presentaron en septiembre (1 153 larvas/1 000 m3) y en noviembre (1 363 larvas/1 000 m3) y en el fondo se obtuvieron en octubre (1 521 larvas/1 000 m3) (Fig. 3c). En la estación cuatro se detectaron dos pulsos máximos en la superficie, el primero fue de 1 092 larvas/1 000 m3 (septiembre) y el segundo de 2 262 larvas/1 000 m3 (octubre). En el fondo se presentaron tres máximos: 568 (julio), 957 (octubre) y 484 larvas/1 000 m3 (diciembre) (Fig. 3d).
En la estación uno se presentaron las más altas densidades; sin embargo, las diferencias en abundancia entre las 4 estaciones de muestreo no fueron significativas (Wilson p>0.05). Por otro lado, la variación de densidad durante el ciclo de estudio presentó diferencias altamente significativas (Wilson p<0.05), quedando divididos en tres grupos (Conover p<0.05): 1) los meses de densidad escasa (febrero-abril), 2) un período de transición (mayo) y 3) los meses en los que se recolectaron filosomas (junio a diciembre).
Las relaciones entre la densidad de filosomas y la temperatura y entre la densidad y la salinidad no fueron significativas (r= 0.232 y 0.125, respectivamente).
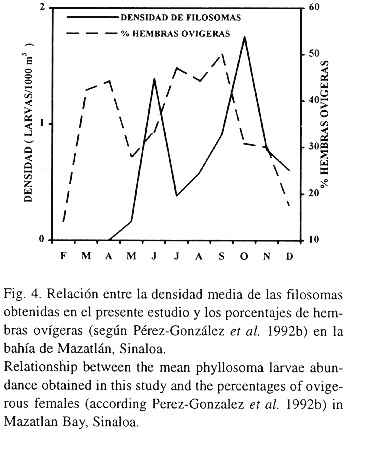
La comparación gráfica entre los porcentajes de las hembras ovígeras obtenidas en esta área por Pérez-González et al. (1992b) y la densidad media mensual de las filosomas obtenidas en este estudio, mostró un desfasamiento aproximado de 8 a 9 semanas, entre los máximos y mínimos porcentajes de las hembras con huevos y de las densidades máximas y mínimas de las filosomas (Fig. 4).
Discusión
Las variaciones de la temperatura y de la salinidad fueron las típicas de esta región (Anónimo, 1974). Se diferenciaron dos épocas climáticas, con un período de transición: la época de secas comprendida entre febrero y junio (salinidades más "altas"), el período de lluvias de julio a octubre (salinidades más "bajas") y los meses de transición entre ambas (noviembre y diciembre con salinidades intermedias). Villalba (1989) encontró este mismo comportamiento y señala que los cambios climáticos influyen en las características de los cuerpos de agua de la zona.
Según las características morfológicas descritas por Johnson y Knight (1966), las filosomas recolectadas correspondieron al estadío I de desarrollo. En la estación uno se registraron las densidades mayores, a pesar de que esta zona no es una área importante de captura comercial. Las zonas más cercanas de pesca son las islas Cardones, Chivos, el cerro del Crestón y las rocas Hermanos y las más alejadas son las Tres Islas (Fig. 1). Sin embargo, es importante señalar que las áreas adyacentes, aunque no muy productivas, son zonas pesqueras y que esta estación está altamente influenciada por las corrientes litoral y de marea (Peraza-Vizcarra 1986, Cabrera-Dueñas 1988). Johnson (1960) menciona que las filosomas en sus primeros estadíos son más abundantes y se localizan en las áreas habitadas por adultos. Probablemente, las filosomas encontradas en esta estación provengan de las áreas de pesca de adultos y fueron transportadas hacia esta zona por el efecto de las corrientes de marea. Cabrera-Dueñas (1988) indica que las aguas que penetran al sistema de Urías durante las primeras horas de la pleamar, provienen del este (isla Cardones) y del sur, entrando directamente al estero (estación uno). Cuando la pleamar llega a su punto máximo, el flujo disminuye y cambia su dirección hacia las Tres Islas. En las primeras horas de la bajamar, el agua sale del estero con dirección al sur y al este y, en el punto mínimo de reflujo, la velocidad disminuye y las aguas de la Tres Islas son arrastradas hacia el sur y el este. Peraza-Vizcarra (1986) informa que en invierno las olas más fuertes provienen del noroeste y del norte, produciendo una deriva litoral hacia el sur y, durante el verano, el oleaje procede del sur y del sureste originando una deriva litoral hacia el norte.
Sheard (1949) y Smith (1958) señalan que durante los primeros estadíos de desarrollo, la distribución de las filosomas de P. longipes (A. Milne Edwards, 1868) en Australia y P. argus en Florida, depende de las corrientes superficiales. En el banco de Campeche el transporte de las larvas hacia la porción más occidental del golfo de México, es producido por las corrientes superficiales y por el impulso del viento, principalmente durante la noche (Olvera-Limas y Ordoñez-Alcalá 1988, Torrijos-López 1989).
Durante este estudio, la menor densidad de filosomas se obtuvo en la estación dos. Esto posiblemente se debió a que las larvas fueron transportadas hacia el interior del sistema estuarino (estación uno) o hacia el norte de la bahía, debido a la circulación de las masas de agua a la que está sometida. Se han detectado aguas provenientes del sistema de Urías a dos kilómetros de distancia (Anónimo 1974).
En las estaciones tres y cuatro las densidades fueron muy bajas con respecto a la uno, pero es importante señalar que son zonas más abiertas y mayormente influenciadas por las corrientes propias de la bahía y las del golfo de California, originando una mayor dispersión de las filosomas en esas áreas. Phillips y McWilliam (1986) indican que las corrientes superficiales originadas por el impulso del viento sobre los primeros centímetros del agua, pueden transportar a las larvas en una dirección opuesta a la circulación general de la costa a más de 100 km. Por otro lado, Johnson (1971, 1974) encontró larvas de P. gracilis en áreas distantes hasta de 2 325 km de la costa, al oeste de las islas Galápagos. Estas zonas se encuentran bajo la influencia de las corrientes del Pacífico Central y del sector occidental del Pacífico Oriental, lo que permite una amplia dispersión de las larvas.
No se presentaron diferencias significativas entre la abundancia de filosomas y las diferentes estaciones de muestreo. Posiblemente, esto se debió a que las recolectas se realizaron en áreas de poca profundidad (entre 6 y 19 m) y muy próximas a la costa (entre cero y 2 km de distancia). Los resultados obtenidos por Johnson (1960), desde Punta Eugenia hasta la parte final de la península de la Baja California, muestran que en áreas costeras con profundidades de entre 0 y 140 m, no existen diferencias apreciables en la densidad de las filosomas. Además, este mismo autor informa que la escasez de las larvas en los primeros estadíos indica, en parte, una tendencia de la larva a buscar lugares profundos, los cuales son de difícil acceso para realizar muestreos. Ritz (1972) encontró que las filosomas presentan una migración vertical activa espacio-temporal en la columna de agua, debido a que responden rápidamente a los cambios de iluminación diurna y nocturna y señala que el comportamiento de las larvas ocasiona que no presenten una distribución uniforme durante las noches, haciendo que se sitúen en niveles más profundos conforme avanza su desarrollo.
Por otro lado, los patrones de variación temporal de la densidad de larvas fueron iguales en todas las estaciones de muestreo. Probablemente a esto se debieron las diferencias mensuales altamente significativas, por lo que el ciclo anual se dividió en tres períodos. A mediados de la primavera se observaron las primeras larvas en toda la zona y continuaron apareciendo durante el verano y parte del otoño. Las más altas densidades se presentaron durante junio (1 390 larvas/1 000 m3) y octubre (1 760 larvas/1 000 m3) (Fig. 4). Johnson (1956) encontró una máxima densidad de 488 larvas/1 000 m3 en las costas del sureste de California, Estados Unidos, y sugiere que el estadío I puede ser bastante abundante en zonas costeras, principalmente frente a Baja California. Tlahuel-Vargas (1992) encontró resultados similares a los de este estudio, con abundancias mayores durante julio, septiembre y octubre y menores el resto del año.
En Baja California, la presencia de filosomas de P. interruptus y P. gracilis es más intensa en verano y en otoño, con una abundancia mayor en agosto y en septiembre; en ocasiones se presentan cantidades similares en julio y octubre y raras veces en junio, noviembre y diciembre (Johnson 1960). En el caso de P. interruptus, el número máximo de hembras ovígeras se presenta a fines de junio y si se considera que el tiempo de incubación de los huevos es de 9 a 10 semanas, la mayor abundancia de filosomas, en el estadio I, podría ocurrir a fines de agosto (Allen 1916).
Las especies de langostas de esta zona, P. inflatus y P. gracilis, presentan reproducción continua casi todo el año, con una reducción aparente en invierno y las épocas de mayor porcentaje de hembras con huevecillos son primavera y verano (Pérez-González et al. 1992b). Probablemente el tiempo de incubación de los huevos en estas especies sea también entre 9 y 10 semanas y a esto se debe el desfasamiento que se presenta entre el porcentaje de hembras ovígeras y la densidad de filosomas (Fig. 4).
Los índices de correlación entre la densidad de filosomas y la temperatura y la salinidad fueron tan bajos, que no se podrían considerar como las variables que determinen directamente la aparición de las larvas, sino que la relación es de manera indirecta, sobre todo con la temperatura, ya que es la que induce al apareamiento y reproducción de los adultos. Este comportamiento debe ser diferente según la especie y/o la zona en que habiten. Sin embargo, en experimentos de laboratorio y observaciones de las filosomas en su hábitat natural, se ha encontrado que larvas de otras especies muestran una preferencia por ciertos intervalos de temperatura y salinidad (Robertson 1971, Kittaka 1994, Rothlisberg et al. 1994). Por otro lado, Alfonso et al. (1995) observaron que las filosomas de P. argus presentaron fototropismo negativo, relacionado con la temperatura, y que las máximas densidades se encontraron por encima de la isoterma y termoclina de los 25.0 y 26.0°C, respectivamente, en el sur del golfo de Batabanó, Cuba.
Phillips y Sastry (1980) afirman que la distribución y la abundancia de las larvas está determinada por la distribución de las hembras ovígeras, duración de la vida pelágica, velocidad y dirección de las corrientes en la profundidad donde se encuentran, respuesta de los diferentes estadíos larvarios a la temperatura, salinidad, baja presión hidrostática y por los intervalos de mortalidad específica de cada estadío.
Agradecimientos
Se agradece a Arturo Nuñez y Luis M. Valadez por su colaboración en la obtención de las muestras. A Fernando González y Roberto Cortés por las facilidades brindadas en las instalaciones del Instituto de Ciencias del Mar y Limnología-Estación Mazatlán, U.N.A.M., en particular el Laboratorio de Plancton. A José Peláez H. por la traducción del resumen. Esta investigación fue financiada por la Dirección General de Investigación Científica y Superación Académica-S.E.P. (Proyecto: 89-01-0274).
Resumen
Se analizó la distribución espacio-temporal de las larvas filosomas de las langostas Panulirus inflatus y P. gracilis y su relación con la temperatura y salinidad, en el sureste del golfo de California, de febrero a diciembre de 1989. Tres estaciones fueron establecidas en la bahía de Mazatlán y una en la boca del sistema de Urías (estación uno). Se obtuvieron 208 muestras (104 de superficie y 104 de fondo) de arrastres horizontales de plancton. Simultáneamente se midieron la temperatura y salinidad del agua. Las larvas fueron más abundantes en la estación uno, con una densidad promedio total de 1 206 y 1 196 larvas/1 000 m3 en superficie y fondo, respectivamente. La temperatura varió de 17.9, en febrero, a 30.2°C, en agosto, y la salinidad de 34.1, en agosto, a 35.000/00, en mayo. La abundancia de larvas no presentó relación con los cambios de temperatura y salinidad (p>0.05); mientras que con la conducta reproductiva de las especies de langosta de esta zona estuvo estrechamente relacionada. La distribución espacial de las filosomas está sujeta a las corrientes y a los ciclos de marea de esta región.
Referencias
Alfonso, I., M. P. Frias, J. Baisre & B. Hernández. 1995. Distribución vertical de filosomas de Panulirus argus y su relación con algunos factores hidroclimáticos al sur del golfo de Batabanó, Cuba. Revista Cubana de Investigaciones Pesqueras 19: 3-9. [ Links ]
Anónimo. 1974. Estudio geográfico de la región de Mazatlán. Dirección General de Oceanografía y Señalamiento Marítimo. Secretaría de Marina, México, D. F. 167 p. [ Links ]
Anónimo. 1980. Estudio integral de la bahía de Mazatlán, Sinaloa. Primer informe parcial del proyecto PCMANAL 790001. Centro de Ciencias del Mar y Limnología, Estación de Investigaciones Marinas, Universidad Nacional Autónoma de México. Mazatlán, México. 22 p. [ Links ]
Allen, B. K. 1916. Notes on the spiny lobster (Panulirus interruptus) of the California Coast. Univ. Calif. Publ. Zool. 16: 139-152. [ Links ]
Báez, P. 1983. Larvas phyllosoma y puerulos de la langosta verde Panulirus gracilis Streets 1871 procedentes de la expedición Costa Rica 1973 (Crustacea: Decapoda: Palinuridae). Rev. Biol. Mar. Valparaíso 19: 79-111. [ Links ]
Buesa, R. J. 1969. Langosta común Panulirus argus (Latreille). Hacia nuevos objetivos en su investigación. Mar y Pesca. Inst. Nal. Pesca, Cuba 46: 4-11. [ Links ]
Cabrera-Dueñas, H. 1988. Modelación hidrodinámica numérica del Puerto de Mazatlán y sus proximidades. Tesis de Licenciatura, Universidad de Colima, Colima, México. [ Links ]
Conover, W. J. 1980. Practical Nonparametric Statistics. Wiley, New York. 493 p. [ Links ]
Chittleborough, R. G, & L. R. Thomas. 1969. Larval ecology of the western Australian marine crayfish, with notes upon other palinurid larvae from the eastern Indian Ocean. Aust. J. Mar. Freshwat. Res. 20: 199-223. [ Links ]
Flores-Campaña, L. M., R. Pérez-González & A. Nuñez-Pasten. 1993. Biología y pesquería de las langostas Panulirus inflatus (Bouvier) y P. gracilis Streets en la costa sureste del Golfo de California, p. 113-121. In González-Cano, J. M. & R. Cruz-Izquierdo (eds.). La utilización de refugios artificiales en las pesquerías de langosta: sus implicaciones en la dinámica y manejo del recurso. Instituto Nacional de la Pesca, Secretaría de Pesca, Centro de Investigaciones Pesqueras de Cuba. Isla Mujeres, México. [ Links ]
Holthuis, L. B. 1991. Marine lobsters of the world. An annotated and illustrated catalogue of species of interest to fisheries known to date. FAO Fisheries Synopsis 13: 1-292. [ Links ]
Johnson, M. W. 1956. The larval development of the California spiny lobster Panulirus interruptus (Randall), with notes on P. gracilis Streets. Proc. Cal. Acad. Sci. 29: 1-19. [ Links ]
Johnson, M. W. 1960. Production and distribution of larvae of the spiny lobster Panulirus interruptus (Randall) with records on P. gracilis Streets. Bull. Scripps Inst. Oceanogr. 7: 413-462. [ Links ]
Johnson, M. W. 1971. The palinurid and scyllarid lobster larvae of the tropical eastern Pacific and their distribution as related to the prevailing hydrography. Bull. Scripps Inst. Oceanogr. 19: 1-36. [ Links ]
Johnson, M. W. 1974. On the dispersal of lobster larvae into the east Pacific barrier (Decapoda, Palinuridae). Fish. Bull. 72: 639-647. [ Links ]
Johnson, M. W. y M. Knight. 1966. The phyllosoma larvae of the spiny lobster Panulirus inflatus (Bouvier). Crustaceana 10: 31-47. [ Links ]
Katz, C. H., J. S. Cobb & M. Spaulding. 1994. Larval behavior, hydrodynamic transport, and potential offshore recruitment in the american lobster, Homarus americanus. Mar. Ecol. Progr. Ser. 103: 265-273. [ Links ]
Kittaka, J. 1994. Culture of phyllosomas of spiny lobster and its application to studies of larval recruitment and aquaculture. Crustaceana 66: 25-270. [ Links ]
Olvera-Limas, R. M. & L. Ordoñez-Alcalá. 1988. Distribución, abundancia relativa y desarrollo larvario de langostas Panulirus argus y Scyllarus americanus en la Zona Económica Exclusiva del Golfo de México y Mar Caribe. Ciencia Pesquera. Inst. Nal. Pesca. Sría. Pesca. México 6: 7-31. [ Links ]
Pearce, A. F. & B. F. Phillips. 1988. ENSO event, the Leeuwin Current and larval recruitment of the western rock lobster. J. Cons. Int. Explor. Mer. 45: 13-21. [ Links ]
Peraza-Vizcarra, R. 1986. Las playas, la deriva litoral de la arena en las playas de la costa sur de Sinaloa. Ciencias del Mar 8: 50-52. [ Links ]
Pérez-González, R. , L. M. Flores-Campaña & A. Nuñez-Pasten. 1992a. Análisis de la distribución de tallas, captura y esfuerzo en la pesquería de las langostas Panulirus inflatus (Bouvier, 1895) y P. gracilis Streets, 1871 (Decapoda: Palinuridae) en las costas de Sinaloa, México. Proc. San Diego Soc. Nat. His. 15: 1-5. [ Links ]
Pérez-González, R. , L. M. Flores-Campaña, A. Nuñez-Pasten & A. A. Ortega-Salas. 1992b. Algunos aspectos de la reproducción en Panulirus inflatus (Bouvier) y P. gracilis Streets (Decapoda: Palinuridae) en el sureste del golfo de California, México. Inv. Mar. CICIMAR 7: 25-33. [ Links ]
Phillips, B. F. 1981. The circulation of the south eastern Indian ocean and the planktonic life of the western rock lobster. Oceanogr. Mar. Ann. Rev. 19: 11-39. [ Links ]
Phillips, B. F. & P. S. McWilliam. 1986. The pelagic phase of spiny lobster development. Can. J. Fish. Aquat. Sci. 43: 2153-2163. [ Links ]
Phillips, B. F. & A. N. Sastry. 1980. Larval ecology, p. 11-57. In J. Stanley Cobb & B. F. Phillips (eds.). The biology and management of lobsters, Vol. II. Academic Press, New York. [ Links ]
Pringle, J. D. 1986. California spiny lobster (Panulirus interruptus) larval retention and recruitment: a review and synthesis. Can. J. Fish. Aquat. Sci. 43: 2142-2152. [ Links ]
Ritz, D. A. 1972. Behavioural response to light intensity of the newly hatched phyllosoma larvae of Panulirus longipes cygnus George (Crustacea: Decapoda: Palinuridae). J. Exp. Mar. Biol. Ecol. 10: 105-114. [ Links ]
Robertson, P. B. 1971. The larvae and postlarva of the scyllarid lobster Scyllarus depressus (Smith). Bull. Mar. Sci. 21: 841-865. [ Links ]
Rothlisberg, P. C., C. J. Jackson, B. F. Phillips & P. S. McWilliam. 1994. Distribution and abundance of scyllarid and palinurid lobster larvae in the Gulf of Carpentaria, Australia. Aust. J. Mar. Freshwat. Res. 45. 337-349. [ Links ]
Sheard, K. 1949. The marine crayfishes (spiny lobsters), family Palinuridae of western Australia. Aust., CSIRO Bull. 247: 1-45. [ Links ]
Smith, F. G. W. 1958. The spiny lobster industry of Florida. Fla. Board. Conserv. Ed. Serv. 11: 1-36. [ Links ]
Tlahuel-Vargas, N. 1992. Abundancia y variación nictemeral de larvas filosomas (Panulirus sp) (1989-1990) en la boca del Puerto de Mazatlán, Sinaloa. México. Tesis de Licenciatura, Universidad Autónoma de Sinaloa, Mazatlán, México. [ Links ]
Torrijos-López, J. 1989. Distribución y abundancia de los estadíos larvales de las langostas Panulirus argus y Scyllarus americanus (Crustacea, Decapoda, Palinuridae) en la Zona Económica Exclusiva del Golfo de México, primavera-verano de 1986 y 1987. Tesis de Licenciatura, Universidad Nacional Autónoma de México, México, D. F., México. [ Links ]
Villalba, L. A. 1989. Descripción general del estero de Urías, Mazatlán, Sinaloa. Ciencias del Mar 8: 32-37. [ Links ]
1 Facultad de Ciencias del Mar, Universidad Autónoma de Sinaloa, Apdo. Postal 610, Mazatlán, Sinaloa, México. Tel./fax (69) 82-86-56, lieb@facimar.maz.uasnet.mx

