Las diatomeas centrales del género Coscinodiscus Ehrenberg pertenecen a un grupo muy grande e importante del fitoplancton marino. Algunas especies del género son indicadoras de condiciones marinas ricas en nutrientes o de surgencias (Smayda 1975, Estrada y Blasco 1979 y Marshall y Cohn 1987), o se usan en experimentos fisiológicos y de reproducción (Schmid 1986, 1990, 1995 y Nagai y Manabe 1994), mientras que otras se emplean en estudios de sedimentología y bioestratigrafía, debido a que varias especies aparecen en períodos geológicos bien definidos (Sancetta y Silvestri 1984). Las especies de Coscinodiscus son también importantes en estudios de evolución, pues son consideradas como ancestros clásicos de otros grupos de diatomeas centrales (Simonsen 1979). Recientemente se han elaborado estudios morfológicos con gran detalle de algunas especies de Coscinodiscus (Brooks 1975 a, b, c, Hasle y Sims 1986 y Makarova 1993), y otros autores han contribuido a clarificar la taxonomía del género (Fryxell 1978 y Fryxell y Ashworth 1988). La identificación a nivel de especie es una tarea difícil, ya que por muchos años los caracteres taxonómicos para separar especies fueron ambiguos, como el arreglo y medidas de las areolas, presencia o ausencia de roseta central, y la presencia y posición de "apículos marginales" ("marginal apiculi") (Hasle y Lange 1992). Sin embargo, algunos otros caracteres son considerados diagnósticos e importantes actualmente, como la presencia y forma de los dos macroprocesos (macrorrimopórtulas), así como la distancia entre ellos, la distancia entre los procesos pequeños (microrrimopórtulas), y la ausencia o presencia de procesos en la cara valvar. Las bandas del cíngulo de algunas especies pueden adicionalmente ayudar en la identificación (Hasle y Lange 1992). 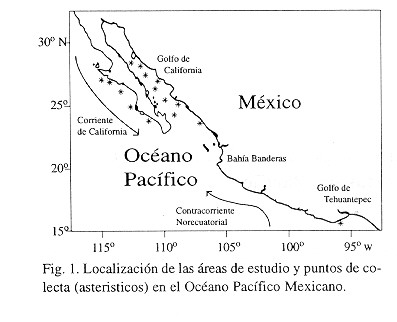
Aunque existen trabajos floríticos de fitoplancton en el Pacífico mexicano, no existe una revisión sobre morfología y taxonomía del género Coscinodiscus, especialmente usando técnicas modernas como el microscopio electrónico. En el Golfo de México se estudiaron tres especies del género (Moreno y Licea 1994), mientras que en una obra sobre diatomeas del Golfo de California (Moreno et al. 1997) se incluyen 16 taxones (12 especies y cuatro variedades). En este trabajo se estudió material de costas del Pacífico mexicano para revisar las especies de Coscinodiscus, tomando en cuenta observaciones en microscopía de luz y electrónica, así como los caracteres taxonómicos de valor.
Materiales y Métodos
El material analizado corresponde a muestras procedentes de costas del Pacífico mexicano, incluyendo costas occidentales de Baja California, Golfo de California y Golfo de Tehuantepec. Se usaron redes de fitoplancton (30 m y 54 µm de luz de malla), durante cinco cruceros (período 1982-1986) y muestras aisladas (Fig. 1). Todas las muestras se preservaron en formaldehído con una concentración final de 4%.
El material se analizó en microscopio de luz (ML), utilizando preparaciones temporales o permanentes, ya sea libres de sales y fijadores o limpio de contenido orgánico (Hasle 1978). Para el estudio con microscopio electrónico de barrido (MEB) se usó material enjuagado o limpio, colocado en dispositivos de aluminio, secado al aire y metalizado con oro. Las observaciones con microscopio electrónico de transmisión (MET) se hicieron únicamente con material limpio, una vez que se colocó sobre rejillas preparadas con Formvar y se secó al aire.La terminología que se sigue es la propuesta por Anónimo (1975), Stosch (1975) y Ross etal. (1979), y las sugerencias de Brooks (1975 a, b, c), Stosch (1980), Fryxell y Ashworth (1988,) y Hasle y Lange (1992).
Resultados
Se identificaron y estudiaron seis especies de Coscinodiscus. Se anotan las observaciones de microscopía (ML, MEB, MET) para cada especie, referencias, datos morfométricos y notas de su distribución local y general. Se sigue un arreglo alfabético. Género Coscinodiscus Ehrenberg
Coscinodiscus asteromphalus Ehrenberg
Hustedt 1930, p. 453, figs. 250 a?c; Cleve?Euler 1951, p. 67, figs. 103?105 d; Brooks 1975a, p. 15, pls. I-VIII; Fryxell y Ashworth 1988, p. 363, figs. 2, 7?8, 14, 18, 26; Hasle y Lange 1992, p. 42, figs. 1-14. Descripción- Las valvas son circulares, ligeramente convexas (Figs. 2, 4), más o menos aplanadas o ligeramente deprimidas en el centro (Fig. 4). Diámetro 115-210 µm. En el centro el número de areolas es de 3-5/10 µm, mientras que en el margen es de 4-6/10 µm. Se presenta una roseta central, formada por 5-9 areolas de mayor tamaño (Fig. 3). También se observaron filas radiales primarias y secundarias de areolas (Fig. 3). Las areolas son hexagonales en su mayoría, con algunas pentagonales al inicio de una fila secundaria. En LM, las cribra aparecen como puntuaciones en las areolas, mientras que en MEB para cada cribrum se nota un patrón de 2-5 poros pequeños centrales, rodeados por un anillo de poros más grandes y alargados (Figs 5, 6); cada cribrum se separa de otro vecino por poros alargados de mayor tamaño. El margen de la valva no muestra estructuras especializadas (Fig. 6). Las microrrimopórtulas se disponen en un anillo localizado a 2-5 areolas del margen valvar. Estos procesos son ligeramente más anchos en sus ápices. Hay dos macrorrimopórtulas casi marginales, asimétricas y separadas por ángulos de 134-147°.
Comentario- Las elementos principales de variación morfológica, en comparación con datos de la literatura son el número de areolas en la roseta central, el ángulo de separación de las dos macrorrimopórtulas y el arreglo de poros que forman cada cribrum. 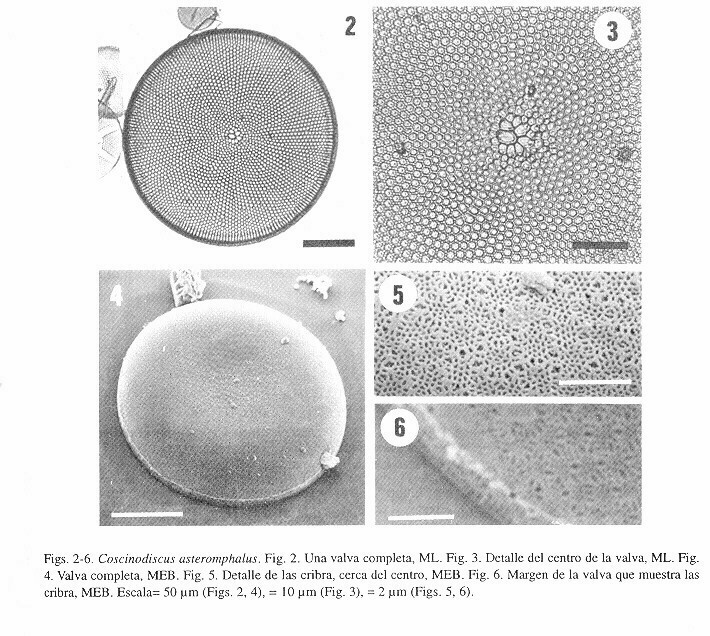
Makarova (1993) mencionó la presencia de rimopórtulas en la cara valvar de C. asteromphalus, mientras que Hasle y Lange (1992, Tabla 1) no encontraron procesos en la cara valvar. Distribución- Es una especie común, aunque no abundante en el Golfo de California y las costas de Baja California. Posiblemente cosmopolita (en cuatro océanos, incluyendo el Antártico) (Hasle y Lange 1992). Coscinodiscus centralis Ehrenberg Descripción- Valvas circulares y ligera o fuertemente cóncavas, deprimidas en el centro (Figs 7, 8, 10, 11). Diámetro 81-227 µm. Las areolas muestran un arreglo radial, partiendo del centro y tienen forma hexagonal (Fig. 9). La densidad areolar es de 3-5 areolas/10 µm en el centro y 3-6 en los márgenes. Existe una roseta central formada por 6-9 areolas de mayor tamaño (Figs 9-11). Cerca del márgen valvar hay un anillo de microrrimopórtulas. Estas son relativamente anchas en la base, pero son más delgadas y aplanadas cerca del ápice. Las macrorrimopórtulas, también cerca del margen, están separadas por un ángulo de 139-154° (Figs. 12, 13). En una vista de la célula completa, las aperturas externas de estos macroprocesos están opuestas (Fig. 12). El cíngulo muestra la presencia de una valvocópula y al menos dos bandas (Figs. 10-12). Comentario- También en esta especie el número de areolas en la roseta central, la separación de las macrorrimopórtulas y la densidad areolar del centro y los margenes son los caracteres con mayor variación.
Existe una estrecha relación entre las especies C. centralis y C. asteromphalus, ambas comparten características comunes muy evidentes como la presencia de la roseta central.
Distribución- Especie de amplia distribución en el Golfo de California y las costas de Baja California. Aparentemente cosmopolita en todos los océanos (Hasle y Lange 1992). 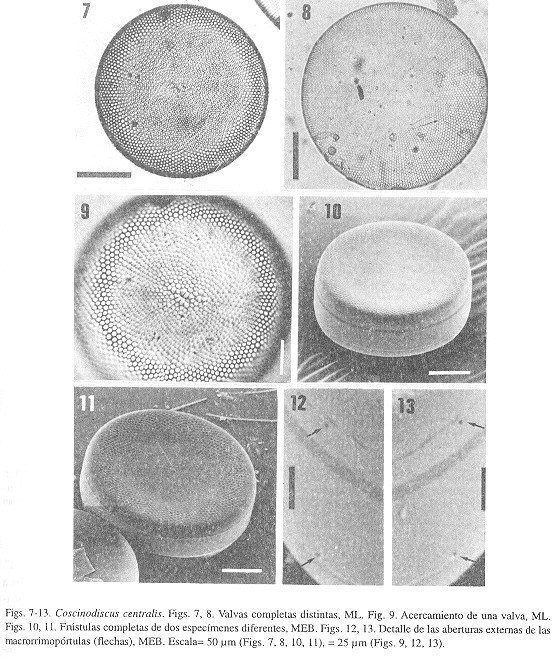
Coscinodiscus concinnus Smith
Hustedt 1930, p. 441, figs. 241 a?e; Cupp 1943, p. 58, figs. 22 a?e; Cleve?Euler 1951, p. 68, figs. 108 a?f; Boalch 1971, p. 685, pl. 2, figs. A?F; Hasle y Lange 1992, p. 50, figs. 31-48. Descripción- La frústula completa tiene forma de tambor, las valvas son circulares y ligeramente convexas (Fig. 14). Diámetro 132-280 µm. El arreglo de las areolas es radial, mientras que la densidad areolar es de 5-8 areolas/10 µm en el centro y 6-9 areolas/10 µm en el margen. La roseta central es muy característica ya que aparece como una zona hialina (Fig. 15) o una área granular y está formada por 9-21 areolas; algunos especímenes presentan poros en esta estructura. La parte externa, como en todas las especies estudiadas, muestra los cribra. Cada cribrum consiste de varios poros centrales, rodeados de poros alargados y de mayor tamaño (Figs 16, 17). Existen 1-3 microrrimopórtulas/10 µm entre la cara valvar y el manto, dispuestas en un anillo periférico (Fig. 18). Tienen forma tubular, aunque un poco más anchas en el ápice (Fig. 18). Las dos macrorrimopórtulas, cercanas al margen, están separadas por un ángulo de 149-155°. El cíngulo presenta tres bandas, la cópula es relativamente delgada, mientras que la pleura es mucho más delgada. Distribución- Presente en el Golfo de California y costas de Baja California. Cosmopolita en todos los océanos, excepto el Antártico (Hasle y Lange 1992). 
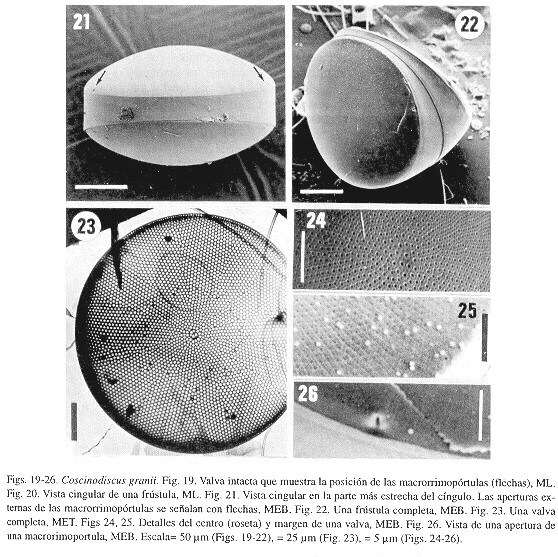
Coscinodiscus granii Gough
Descripción- Las células son discoides en vista valvar (Fig. 19) y cuneiformes en vista cingular (Figs 20-22). Las valvas son circulares y totalmente asimétricas, con un lado más alto que el otro (Fig. 19). Diámetro 155-178 µm. El centro, ligeramente deprimido, lo ocupa una roseta central, formada por 5-6 areolas de mayor tamaño (Figs 23, 24). La areolación sigue un patrón radial, con 7 areolas/10 µm en el centro y 8-9/10 µm en los márgenes. Son notables las filas primarias y secundarias de areolas (Fig. 23). Cada cribrum consiste de un gran poro, rodeado por 5-7 poros más pequeños (Fig. 24). Las áreas subsidiarias (según Brooks, 1975c) son evidentes (Figs 24, 25). Hay un anillo de microrrimopórtulas marginales, separadas por 5-8 areolas. Las dos macrorrimopórtulas están separadas por un ángulo de 135° (131-138°) (Fig. 21). Su localización se muestra por indentaciones pronunciadas (Fig. 19) y externamente muestran apertura marginales grandes y muy conspicuas (Fig. 26). Dichas aperturas están opuestas en las valvas de una misma célula (Fig. 22). Se aprecian dos bandas del cíngulo, aunque algunos especímenes muestran solo una. Estas bandas son típicamente asimétricas: un lado es considerablemente más alto (ancho) que el otro (Figs 21, 22).
Comentario- Esta especie se reconoce fácilmente debido a su forma característica. Las observaciones hechas aquí son consistentes con estudios previos (e.g. Hasle y Lange 1992); sin embargo, la depresión que se señala en el centro de la valva no se ha mencionado con anterioridad. El número de bandas del cíngulo en C. granii es probablemente dos, aunque Brooks (1975c) encontró una tercera ocasionalmente.
Holmes y Reimann (1966) discutieron la identidad de C. granii y propusieron que podría ser considerada como una variedad de C. concinnus. Varios argumentos son contrarios a esta idea (Hasle y Lange 1992); aquí se considera una especie distinta.
Distribución-Coscinodiscusgranii fue encontrada en los Golfos de California y Tehuantepec. Especie considerada cosmopolita en los cinco océanos (Hasle y Lange 1992).
Coscinodiscus radiatus Ehrenberg
Hustedt 1930, p. 420, figs. 225 a?b; Cupp 1943, p. 56, figs. 20 a?d; Cleve?Euler 1951, p. 65, figs. 97 a?b; Hasle y Sims 1986, p. 310, figs. 8?32; Sancetta 1987, p. 234, figs. 1?10; Fryxell y Ashworth 1988, p. 363, figs. 3, 9?10, 15, 19, 27; Moreno y Licea 1994, p. 117, figs. 4, 22-40. Sinónimos: Coscinodiscusperforatus var. cellulosa Grunow
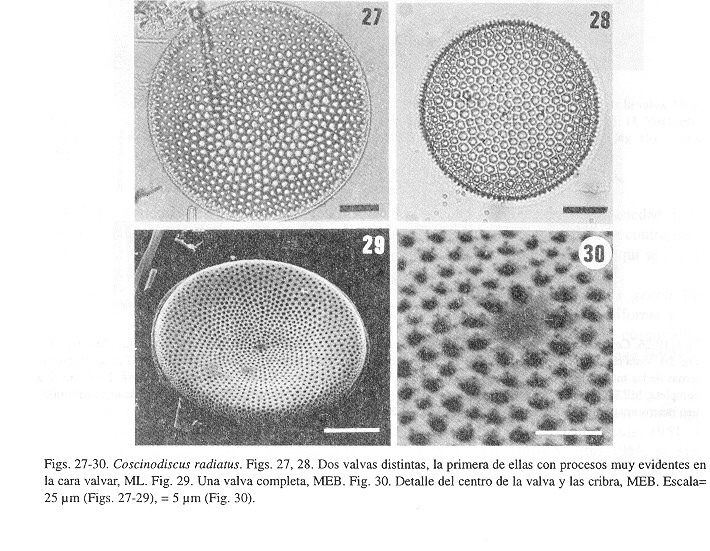
Coscinodiscus perforatus var. pavillardii (Forti) Hustedt
Descripción- Las valvas son circulares y convexas, cóncavas o planas, sin un manto alto conspícuo (Figs 27-29). Diámetro valvar de 46-148 µm. La densidad areolar es de 4-7 areolas/10 µm, en el centro y el margen. Las areolas pueden ser pentagonales, hexagonales o heptagonales (Fig. 28) y muestran filas radiales primarias y secundarias (Fig. 27). La roseta central está formada por 4-8 areolas ligeramente más grandes y a menudo se encuentra una areola central rodeada por 6-7 areolas. En algunos especímenes el área central no presenta areolas (Fig. 30). Los cribra son muy característicos (aunque en el material de MEB no se aprecia bien): un patrón de 5-8 anillos concéntricos de poros, los de mayor talla son los más periféricos y hay áreas hialinas entre los cribra. En el manto, los cribra se sobreponen y se pierde el patrón (Fig. 29). Existen varias rimopórtulas (del tipo micro) en la cara valvar, entre la parte central y el margen, las cuales coinciden con el comienzo de filas secundarias de areolas (Fig. 27). También cerca del margen (separadas del margen por 1 areola) se dispone un anillo de microrrimopórtulas, que en el interior muestran su estructura tubular y están orientadas hacia el centro de la valva. Las dos macrorrimopórtulas están separadas por un ángulo de 132-147°, las cuales se pueden localizar en ML como indentaciones en el margen valvar. No se pudieron estudiar los detalles del cíngulo. Comentario- Existe gran variabilidad morfológica en esta especie, lo que ha sido discutido previamente y ha conducido a la propuesta de colocar los taxones C. perforatus var. cellulosa y C. perforatus var. pavillardii como sinónimos de C. radiatus (Moreno y Licea 1994), siguiendo una sugerencia de Sancetta (1987). Distribución- Especie muy común y ampliamente distribuida en todo el Pacífico mexicano, desde las costas de Baja California y el Golfo de California hasta el Golfo de Tehuantepec.
Coscinodiscus wailesii Gran et Angst
Descripción- Las células tienen forma de tambor y son de una talla relativamente grande (Fig. 34). Las valvas son circulares y relativamente planas, con una ligera depresión en el centro y un manto muy alto (Figs. 31, 33, 34). El diámetro es de 171-365 µm, mientras que el eje pervalvar es de 108-180 µm. En el centro de la valva se presenta una área hialina circular o estrellada (Fig. 32). La areolación es radial, encontrándose filas secundarias de areolas (Fig. 31), las cuales corren a partir de areolas más pequeñas o, en la mayoría de los casos, a partir de una microrrimopórtula de la cara valvar (Fig. 37). El área central se extiende por medio de radios hialinos (o amplia interestría). La densidad areolar es de 6-7 areolas/10 µm en la cara valvar y 5-6 areolae/10 µm en el manto.
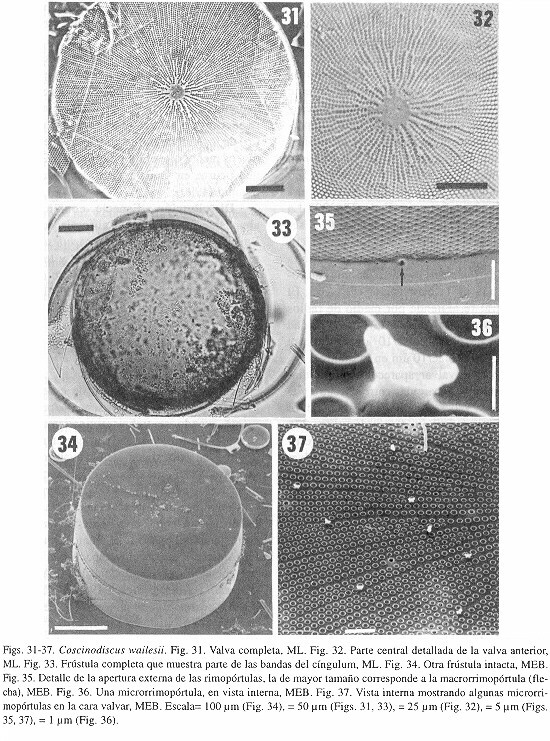
En la cara valvar aparecen rimoportulas (del tipo micro), esparcidas entre el centro y el margen de la valva, generalmente al comienzo de una fila secundaria de areolas (Fig. 37). Adicionalmente hay dos anillos de microrrimopórtulas, uno en el margen de la cara valvar y otro en el margen del manto (2 1/2 procesos/10 µm). En el interior, estos procesos tienen un cuello corto y después se expanden en una estructura curva (Fig. 36). Tanto en la cara valvar como en el manto, los microprocesos se localizan usualmente al final de un radio hialino. Las dos macrorrimopórtulas se encuentran en el manto, entre las micro del anillo marginal, separadas por un ángulo de 135-160°. La apertura externa de los macro es de mayor tamaño que la de las microrrimopórtulas (Fig. 35). El cíngulo, no estudiado en detalle, aparentemente presenta tres bandas, incluyendo una valvocópula (Figs. 33, 34). Comentario- Los caracteres distintivos de esta especie conducen a su relativamente fácil determinación: el tamaño grande de las células, el manto valvar alto, el área central hialina, el tipo de areolación y las rimopórtulas en la cara valvar. Otros estudios (e.g.Schmid 1990) han revelado que la típica área hialina puede transformarse en una roseta central a través de la reproducción vegetativa. La forma de las microrrimopórtulas estudiadas aquí difiere ligeramente de la encontrada por Schmid (1990) y Hasle y Lange (1992), que muestran un cuello más largo. Finalmente se anota que la talla mínima (171 µm de diámetro) de los individuos encontrados en este estudio, es menor que el límite más bajo (280 µm) que ocurre en la naturaleza, establecido por Schmid (1990) Schmid1990 Distribución- Especie común, nunca abundante en el Golfo de California y costas de Baja California. Coscinodiscus wailesii ha sido registrada en áreas subtropicales y templadas de los Océanos Pacífico y Atlántico (Rincé y Paulmier 1986), aunque ausente de los otros tres océanos (Hasle y Lange 1992). Discusión La identificación certera de especies del género Coscinodiscus no es sencilla, particularmente cuando se trabaja rutinariamente con muestras de fitoplancton. Los caracteres taxonómicos de valor siguen siendo evaluados, aunque muchos son muy útiles (como la presencia de procesos en la cara valvar). La forma de la valva también ha sido anotada como un caracter distintivo (como el caso de C. granii), así como la altitud relativa del manto (C. wailesii). Otros caracteres son muy específicos, como la presencia de areola endoquiástica (EAs) en C. alboranii Pavillard (Stosch 1980, 1985).
Es posible que el alto número de taxones de Coscinodiscus (Van Landingham 1968) se vea reducido, al estudiar más especies en detalle y comprobar que muchas son meros sinónimos de otras bien definidas y mejor reconocidas; el caso de C. radiatus es un ejemplo claro (Moreno y Licea 1994).
Respecto a la distribución de las especies encontradas en este trabajo, la mayoría muestra una amplia distribución local y son consideradas cosmopolitas, especialmente Coscinodiscusradiatus, probablemente debido a su extrema variabilidad morfológica, que ha ocasionado que otros taxones sean considerados ahora como sinónimos (e.g. C. perforatus var. cellulosa y C. perforatus var. pavillardii). Coscinodiscuswailesii es la especie que parece más limitada en su distribución (tropical a templada).
Se mencionó la presencia de un número mayor de taxones del género en el Pacífico mexicano (16 en total, de acuerdo a Moreno et al. 1997), aunque no todos han sido estudiados con detalle: la gran mayoría de las ilustraciones de esa obra (Moreno et al. 1997) solo utilizan microscopio de luz y contiene un dibujo (lámina 4, fig. 2) muy deficiente (Coscinodiscusargus Ehrenberg); adicionalmente, la única fotografía de MEB (lámina 18, fig. 7), perteneciente a C. wailessi, deja mucho que desear y no es convincente que se refiere a dicha especie.
Agradecimientos
Varias instituciones y colegas aportaron material para este trabajo, en particular R. Cortés Altamirano (ICMyL, UNAM, Estación Mazatlán). Teté Meave del Castillo (UAM-Iztapalapa) colaboró con diversas facilidades de fotografía. La mayoría de observaciones (especialmente en MEB) fueron hechas en el Depto. de Botánica, de la Universidad de Bristol, Inglaterra.
Resumen
En este trabajo se analizaron muestras de red de fitoplancton procedentes del Pacífico Mexicano, incluyendo los Golfos de California y Tehuantepec, para hacer una revisión actualizada de las especies de diatomeas del género Coscinodiscus. Se encontraron y estudiaron seis especies del género por medio de microscopía de luz y electrónica (principalmente el microscopio electrónico de barrido). Cada descripción de la especie se acompaña de referencias clásicas, discusión de la variabilidad morfológica y se anota la distribución local y general de cada especie. Coscinodiscus radiatus fue la especie más común y ampliamente distribuída, quizás parcialmente debido a que algunos otros taxa se han propuesto como sinónimos de la especie. Se discuten los problemas comunes en la identificación de las especies de Coscinodiscus y se anotan los caracteres taxonómicos de valor: forma de la valva, presencia de procesos valvares, altura relativa del manto y otros caracteres específicos.
Referencias Anónimo. 1975. Proposals for a standardization of diatom terminology and diagnoses. Nova Hedwigia, Beih. 53: 323?354. [ Links ]
Boalch, G.T. 1971. The tipification of the diatom species Coscinodiscusconcinnus WM. Smith and Coscinodiscus granii Gough. J. Mar. Biol. Ass. U.K. 51: 685?695. [
Links ]
Brooks, M. 1975a. Studies on the genus Coscinodiscus I. Light, transmission and scanning electron microscopy of C. concinnus Wm. Smith. Bot. Mar. 18: 1?13. [
Links ]
Brooks, M. 1975b. Studies on the genus Coscinodiscus II. Light, transmission and scanning electron microscopy of C. asteromphalus Ehr. Bot. Mar. 18: 15?27. [
Links ]
Brooks, M. 1975c. Studies on the genus Coscinodiscus III. Light, transmission and scanning electron microscopy of C. granii Gough. Bot. Mar. 18: 29?39. [
Links ]
Cleve?Euler, A. 1951. Die Diatomeen von Schweden und Finnland. K. Svenska Vetensk Akad. Handl, Ser. 4, 4(1), 95?110. [
Links ]
Cupp, E.E. 1943. Marine plankton diatoms of the west coast of North America. Bull. Scripps Inst. Oceanogr. 5: 1?237. [
Links ]
Estrada, M. & D. Blasco. 1979. Two phases of the phytoplankton comunity in the Baja California upwelling. Limnol. Oceanogr. 24: 1065?1080. [ Links ]Fryxell, G.A. 1978. Proposal for the conservation of the diatom Coscinodiscusargus Ehrenberg as the type of the genus. Taxon 27: 122?125. [
Links ]
Fryxell, G.A. & T.K. Ashworth. 1988. The diatom genus Coscinodiscus Ehrenberg: characters having taxonomic value. Bot. Mar. 31: 359?374. [ Links ]
Gran, H.H. & E.C. Angst. 1931. Plankton diatoms of Puget Sound. Pub. Puget Sound Biol. Sta. 7: 417-519. [ Links ]
Hasle, G.R. 1978. Diatoms, p. 136-142. In A. Sournia (ed). Phytoplankton Manual. UNESCO, Paris. [ Links ]
Hasle, G.R. & C.B. Lange. 1992. Morphology and distribution of Coscinodiscus species from the Oslofjord, Norway, and the Skagerrak, North Atlantic. Diatom Res. 7: 37-68. [
Links ]
Hasle, G.R. & P.A. Sims. 1986. The diatom genus Coscinodiscus Ehrenb. C. argus Ehrenb. and C. radiatus Ehrenb. Bot. Mar. 29: 305?318. [
Links ]
Holmes, R.W. & B.E.F. Reimann. 1966. Variation in valve morphology during the life cycle of the marine diatom Coscinodiscus concinnus. Phycologia 5: 233?244. [
Links ]
Hustedt, F. 1930. Die Kieselalgen Deutschlands Österreichs und der Schweiz und der Berüksichtigung der übrigen Länder Europas Sowie der Angrenzenden Meeresgebiete. Die Kryptogamenflora Von Deutschland Österreich und der Schweiz 7 (1). Akad. Verlag., Leipzig. 920 p. [
Links ]
Makarova, I. 1993. Morphology of some species of the genus Coscinodiscus Ehrenberg. Nova Hedwigia, Beih. 106: 185-196. [
Links ]
Marshall, H.G. & M. Cohn. 1987. Phytoplankton composition of the New York Bight and adjacent waters. J. Plankton Res. 9: 267?276. [
Links ]
Moreno, L. & S. Licea. 1994. Morphology of three related Coscinodiscus Ehrenberg taxa from the Southern Gulf of Mexico and coastal North Pacific of Mexico. 11th Diatom Symposium 1990, Mem. Calif. Acad. Sci. 17: 113-127. [ Links ]Moreno, J.L., S. Licea & H. Santoyo, 1996. Diatomeas del Golfo de California. Universidad Autónoma de Baja California Sur, México. 273 p. [
Links ]
Nagai, S. & T. Manabe. 1994. Auxospore formation of a giant diatom, Coscinodiscuswailesii (Bacillariophyceae), in culture. Bull. Plankton Soc. Japan 40: 151-167. [ Links ]
Rincé, Y. & G. Paulmier. 1986. Données nouvelles sur la distribution de la diatomée marine Coscinodiscus wailesii Gran & Angst (Bacillariophyceae). Phycologia 25: 73-79. [ Links ]
Ross, R., E.J. Cox, N.I. Karayeva, D.G. Mann, T.B.B. Paddock, R. Simonsen & P.A. Sims. 1979. An amended terminology for the siliceous components of the diatom cell. Nova Hedwigia, Beih. 64: 511?530. [ Links ]
Sancetta, C. 1987. Three species of Coscinodiscus Ehrenberg from North Pacific sediments examined in the light and scanning electron microscopes. Micropaleontology 33: 230?241. [ Links ]
Sancetta, C. & S. Silvestri. 1984. Diatom stratigraphy of the late Pleistocene (Brunhes) subarctic Pacific. Mar. Micropaleontol. 9: 263?274. [ Links ]
Schmid, A.-M. M. 1986. Wall morphogenesis in Coscinodiscus wailesii Gran et Angst. II. Cytoplasmic events of valve morphogenesis, p. 293-314. In M. Ricard (ed). Proc. 8th Internat. Diatom Symp., Paris, 1984. Koeltz, Koenigstein. [ Links ]
Schmid, A.-M. M. 1990. Intraclonal variation in the valve structure of Coscinodiscuswailesii Gran et Angst. Nova Hedwigia, Beih. 100: 101-119. [ Links ]
Schmid, A.-M. M. 1995. Sexual reproduction in Coscinodiscus granii Gough in culture: a preliminary report. Proc. 13th Internat. Diatom Symp., 1994. Biopress, Bristol. pp. 139-159. [ Links ]
Smayda, T. 1975. Net phytoplankton and the greater than 20 micron phytoplankton size fraction in upwelling waters off Baja California. Fish. Bull. 73: 38-50. [ Links ]
Simonsen, R. 1979. The diatom system: ideas on phylogeny. Bacillaria 2: 1?71. [ Links ]
Stosch, H.A. von. 1975. An amended terminology of the diatom girdle. Nova Hedw., Beih. 53: 1-28. [ Links ]
Stosch, H.A. von. 1980. The 'endochiastic areola', a complex new type of siliceous structure in a diatom. Bacillaria 3: 21-40. [ Links ]
Stosch, H.A. von. 1985. Some marine diatoms from the australian region, specially from Port Phillip Bay and tropical north?eastern Australia. Brunonia 8: 293?348. [ Links ]
Van Landingham, S.A. 1968. Catalogue of the fossil and recent genera and species of diatoms and their synonyms. Part II. Bacteriastrum through Coscinodiscus. J. Cramer, Vaduz. p. 494?1086. [ Links ]
1 Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México (UNAM), Apdo. postal 70-305, Coyoacán, México, D.F. 04510 México.
FAX: +52-5-616 0748
Cornisa: Coscinodiscus del Pacífico Mexicano












 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
