











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.47 no.4 San José dic. 1999
Abstract
A new species of treefrog, Tepuihyla celsae (Hylidae), is described based on 11 specimens from cloud-forests (1250 m) of the Sierra de San Luis, state of Falcón, northwestern Venezuela. The new species is distinguished from its congeners by dorsal pattern dark-brown with some few, small and scattered white spots, upper lip immaculate white, dorsum strongly tuberculated in both sexes, dorsal spicules absent, eye-naris distance longer than horizontal length of eye, fingers lacking webbing, axillary membrane absent. This new Tepuihyla extend the known range of the genus more than 500 km northwesterward, and is the first report of the genus outside the Guyanan masif, adding new evidence to support a close biogeographic affinity among the Sierra de San Luís and the Cordillera de la Costa with the highlands of the Guyanan región.
Key words
Amphibia, Anura, Hylidae, Biogeography, Falcón, Tepuihyla celsae sp. nov., Venezuela, Taxonomy.
Hace treinta años que Rivero (1968) describió a Hyla rodriguezi a partir de ejemplares provenientes de la Sierra de Lema (Gran Sabana, sureste de Venezuela). Desde entonces las relaciones intragenéricas de H. rodriguezi permanecieron ambiguas (Frost 1985, La Marca 1992), hasta que en fecha reciente Duellman & Hoogmoed (1992) transfirieron esta especie al género Osteocephalus. Sin embargo, estos autores debieron modificar la definición de este último género para ajustarlo a ciertas características únicas de H. rodriguezi (e.g. la escasa palmeadura en manos y pies). Casi simultáneamente, Ayarzagüena et al. (1992a) describieron cinco especies de Osteocephalus (O. aecii, O. edelcae, O. galani, O. luteolabris y O. rimarum) que consideraron estrechamente emparentadas con O. rodriguezi, proponiendo reunirlas dentro del grupo de "O. rodriguezi", al que juzgan monofilético con base en la reducida fórmula de palmeadura manual y pedal (Ayarzagüena et al. 1992a: 137). Subsecuentemente, Ayarzagüena et al. (1992b) erigieron el género Tepuihyla para designar a los miembros del grupo de O. rodriguezi al que definieron (entre otras características) por poseer la cabeza tan ancha como larga, carecer o tener rudimentos de membranas en las manos, pies 1/2 a 1/3 palmeados, membrana entre los dos primeros dedos pedales inexistente o muy reducida, espermatozoides con cabeza filiforme y dos flagelos. Más recientemente, Duellman & Yoshpa (1996) adicionan una nueva especie de Tepuihyla, elevando a siete los componentes de este género.
Tiempo atrás el encargado de la colección herpetológica de la Estación Biológica de Rancho Grande (EBRG), informó sobre unas ranas no identificadas provenientes de la Sierra de San Luis, estado Falcón, noroeste de Venezuela. Las caracteríticas morfológicas de estas ranas coincidían con las descritas para el género Tepuihyla, consideradas endémicas de las cimas o alturas medias de los tepuyes en Venezuela y Guyana (Ayarzagüena et al. 1992b, Duellman & Yoshpa 1996). No obstante, los ejemplares falconianos se diferencian claramente de los ejemplares del sur. Como propósito de este trabajo tenemos: (1) reseñar e ilustrar la nueva especie de Tepuihyla y (2) comentar sobre la biogeografía y relaciones intergenéricas del género Tepuihyla.
Materiales y Métodos
La terminología, medidas y caracteres empleados en la descripción son una combinación de las propuestas de Ayarzagüena et al. (1992a) y Heyer et al. (1990); la fórmula que describe la extensión de membrana interdigital pedal sigue la proposición de La Marca (1994). Las medidas se tomaron con un vernier (± 0.1 mm de apreciación) bajo una lupa de disección de la siguiente manera: longitud rostro-cloacal (LRC, desde la punta del hocico hasta la abertura cloacal), largo cabeza (LCB, desde la punta del hocico hasta la comisura de la boca), ancho cabeza (ACB, distancia transversal de la cabeza a nivel de la comisura de la boca), longitud de la tibia (TIB, medida entre la coyuntura tibio-femoral y la coyuntura tibio-tarsal), longitud de la pata (PAT, longitud medida desde el borde posterior del tubérculo metatársico interno hasta el borde externo de la almohadilla del dedo pedal más largo), longitud de la mano (MAN, medida desde el borde posterior del tubérculo palmar hasta el borde externo de la almohadilla del dedo manual más largo), distancia hocico-ojo (DHO, desde la punta del hocico hasta el borde anterior del ojo), distancia ojo-narina (DON, distancia entre el borde anterior del ojo y la abertura narinal), longitud horizontal del ojo (LHO, longitud entre los bordes anterior y posterior del ojo), longitud horizontal del tímpano (LHT, longitud antre los bordes anterior y posterior del tímpano, incluyendo el anillo). La determinación de sexo se hizo por examen directo de las gónadas; como criterio de adultez se consideró la presencia de huevos maduros en las hembras y testículos agrandados y/o hendiduras vocales abiertas en los machos. Los ejemplares estudiados (N = 11) se encuentran depositados en la colección de la Estación Biológica de Rancho Grande, Ministerio del Ambiente y de los Recursos Naturales Renovables, Maracay, Venezuela (EBRG).

Holotipo: EBRG 3424, hembra adulta, proveniente de Cerro Galicia (11°10 N, 69°42 W), Curimagua, Sierra de San Luis, Municipio Miranda, estado Falcón, Venezuela, aproximadamente 1250 m. Recolector: G. Pérez. Fecha: mayo, 1987.
Paratopotipos: EBRG 3423, 3425-26, 3430, 3432 (hembras adultas) y 3422 (hembra inmadura), EBRG 3427-3429 (machos adultos), EBRG 3431 (juvenil, sexo indefinido), todos con los mismos datos que el holotipo.
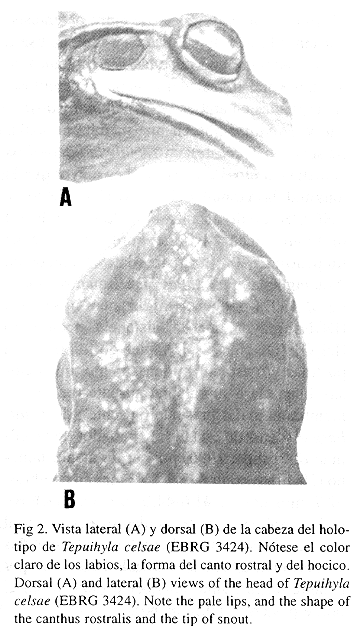
Definición y diagnosis: Esta nueva especie habita en el bosque nublado de la Sierra de San Luis (todos los ejemplares conocidos fueron encontrados a 1250 m de altitud) y se distingue de sus congéneres por la siguiente combinación de características: (1) color dorsal pardo oscuro con unas pocas manchas blancas diminutas y dispersas, (2) labio superior blanco inmaculado, (3) superficie dorsal de los muslos sin barras transversales, (4) hocico corto, semicircular en vista dorsal, (5) talones alcanzan hasta el borde posterior del tímpano cuando estos se extienden hacia adelante, (6) tubérculo subarticular distales del III dedo manual bífido o doble, (7) tubérculo subarticular del IV dedo de la mano dividido, (8) tamaño relativamente grande (longitud rostro-cloacal 0 =52.1 mm en hembras, 0 =42.8 mm en machos), (9) fórmula de la membrana interdigital del pie I0.5-0.0II(1.0-2.0)-1.0III(2.5-3.0)-(2.0-2.5)IV(2.0-3.0)-(2.5-3.0)V, (10) membrana axilar ausente, (11) cabeza ligeramente más ancha que larga, (12) abertura cloacal bordeada ventralmente por dos tubérculos situados lateralmente a la abertura cloacal, grandes, obtusos, con muchos y profundos pliegues, (12) discos terminales de los dedos pedales II y III con surcos antero-ventrales en forma de "m".
Tepuihyla celsae se distingue fácilmente de casi todos sus congéneres por su mayor tamaño (LRC en T. celsae, machos 41.6-44.0 mm, hembras 43.5-56.2; T. aecii, machos 28.1-34 mm, hembra 36.8 mm; T. edelcae, machos 30.0-40.2 mm, hembras 32.0-45.7 mm; T. galani, machos 36.8-40.0 mm, hembra 49.5 mm; T. rimarum, machos 27.0-31.8 mm, hembras 30.0-44.6 mm; T. rodriguesi, machos 27.6-34.7 mm, hembras 35.9-38.1 mm; T. talbergae, macho 32.7 mm, hembra 29.1 mm). Además, se diferencia de T. aecii, T. edelcae, T. galani y T. rimarum (datos tomados de Ayarzagüena et al. 1992a; datos de T. celsae entre paréntesis) por tener la piel dorsal lisa en hembras, lisa u ocasionalmente con pústulas y/o espículas en machos (piel dorsal densamente tuberculosa, sin espículas en ambos sexos); Tepuihyla edelcae y T. galani poseen una membrana axilar (ausente en T. celsae); en Tepuihyla luteolabris la distancia ojo-narina [DON] es mayor que la longitud horizontal del ojo [LHO] (DON < LHO), el tobillo alcanza hasta la región loreal (el tobillo alcanza hasta el borde posterior del tímpano); de T. rodriguezi (datos tomados de Ayarzagüena et al. 1992a; Duellman & Hoogmoed 1992) por tener la distancia ojo-narina [DON] mayor que la distancia hocico-ojo [DHO] (DON < DOH), piel dorsal lisa con pústulas y/o espículas (densamente tuberculosa), línea clara supralabial ausente (presente), membrana axilar presente (ausente); de T. talbergae (datos de esta última tomados de Duellman & Yoshpa, 1996) por ser de menor tamaño (29.1 mm contra 43.5-56.2 mm en hembras, 32.7 mm contra 41.6-44.0 mm en machos), piel dorsal lisa con espículas dispersas en el macho (densamente tuberculosa sin espículas en machos), tobillo se extiende hasta algo por delante de la punta del hocico (sólo hasta el borde posterior del tímpano).
Las especies de Phrynohyas y Osteocephalus muestran una apariencia externa superficialmente similar a la nueva especie de Tepuihyla. Sin embargo T. celsae se distingue de cualquiera de las especies registradas en Venezuela para ambos géneros (La Marca, 1997c), por las ausencia de palmeadura en las manos y entre los dedos pedales I y II.
Descripción del holotipo: Cabeza apenas ligeramente más ancha que larga, corta (32.8% de la longitud rostro cloacal); hocico corto (DON/DHO >> 1), semicircular en vista dorsal, redondeado en vista lateral; región interorbital algo cóncava; longitud horizontal del ojo ligeramente mayor que la distancia ojo-narina; narina 2.7 veces más cerca de la punta del hocico que del borde anterior del ojo; distancia interorbital 1.3 veces mayor que el ancho del párpado superior, casi 1.5 veces mayor que la distancia internarinal; anillo timpánico casi 3/4 de la longitud horizontal del ojo, ambos separados por una distancia igual a 1.5 veces la longitud horizontal del anillo timpánico; anillo timpánico con rebordes gruesos y ligeramente elevados, borde superior cubierto por un pliegue glandular que recorre hasta el hombro; lengua redondeada, tan larga como ancha, entera, aproximadamente 1/5 posterior libre; odontoides vomerianos en dos grupos transversos situados entre y detrás de las coanas; canto rostral redondeado; región loreal moderadamente inclinada, cóncavo. Piel dorsal densamente tuberculosa, glandular; densidad de tubérculos es mayor en la región cefálica (inclusive sobre los párpados) que en el resto del dorso; tubérculos pronunciados; flancos igual que dorso; glándulas parotoides presentes, alargadas, estrechas, situadas algo posteriormente y por encima del tímpano; piel dorsal de los miembros corrugada, apenas con algunos tubérculos; toda la región ventral fuertemente areolada. Pupila horizontal. Abertura cloacal situada por encima de la línea media de los muslos, dirigida ventralmente, cubierta parcialmente por un pliegue dorsal grueso, con bordes algo irregulares; dos protuberancias carnosas, elevadas, arrugadas, situadas lateralmente a la abertura cloacal y ventral al borde del pliegue cloacal, y entre ambas un surco muy profundo que no alcanza el borde ventroposterior del cuerpo. Dientes premaxilares y maxilares presentes, cubiertos por un pliegue labial, mucho más pequeños que los dientes vomerianos.
Miembros delanteros largos, delgados; manos grandes, 30.8% de la longitud rostro-cloacal (LRC); patas pequeñas, delgadas, cortas; pata trasera 36.6% de LRC; fémur 45% de LRC; rudimento de pólex visible externamente; tubérculos supernumerarios presentes, pequeños, redondeados, achatados; tubérculos subarticulares grandes, prominentes (en vista lateral); tubérculos subarticulares bífidos, el penúltimo del dedo III de la mano, y el último del dedo IV de la mano; primer dedo de la mano mucho más corto que el segundo; tamaño relativo de los dedos de la mano I<II<IV<III; dedos de la mano sin membrana interdigital evidente; rebordes laterales ausentes; dedos de la mano con discos redondeados, grandes, 2.0 veces más anchos que las falanges adyacentes; disco mayor en el tercer dedo, ligeramente de menor que la longitud horizontal del tímpano cuando se coloca sobre este; discos con una hendidura profunda alrededor del margen distal; tubérculo metatársico interno mucho más largo que ancho (2.7 veces más largo), margen libre redondeado, protuberante; tubérculo metatársico externo 2.0 veces más pequeño que el tubérculo metatársico externo, ligeramente más largo que ancho, redondeado en vista dorsal, achatado; tubérculos supernumerarios presentes, pequeños, redondeados, achatados; tubérculos subarticulares grandes, redondeados, achatados (en vista dorsal), subcónicos en vista lateral, presentes sólo en la palma del pie; dedos del pie terminan en un disco redondeado grande, 1.9 veces más ancho que la falange adyacente; disco mayor en el tercer dedo del pie, ligeramente menos ancho que la longitud horizontal del tímpano cuando se coloca encima de éste; discos con una hendidura profunda alrededor del margen distal; dedos del pie II y III con una hendidura media y transversal a la anterior, menos profunda, formando una "m"; dedos del pie presentan la membrana interdigital como sigue: I1.0-1.5II2.5-2.0III3.5-3.0IV4.0-4.0V; tamaño relativo de los dedos del pie I<II<III=V<IV; pliegue tarsal ausente; el talón apenas llega hasta el borde posterior del anillo timpánico cuando las patas traseras se estiran hacia adelante.
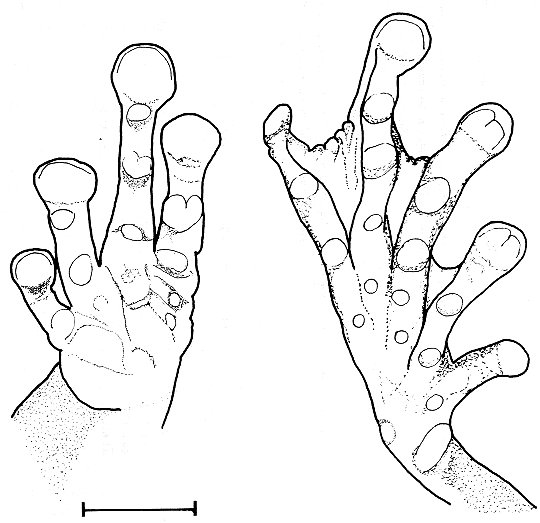
Fig. 3. Dibujo esquemático de la mano (A) y pie (B) en vista ventral, respectivamente, de Tepuihyla celsae (EBRG 3424). Escala = 5.0 mm.
Drawings of the hand (A) and the foot (B) in ventral view of Tepuihyla celsae (EBRG 3424). Scale bar = 5.0 mm.
Coloración en preservativo: Superficie dorsal castaño oscuro, con unas pocas manchas blancas, pequeñas, dispersas, en la cabeza y dorso; cabeza más pálida que el dorso; labio superior blanco inmaculado; labio inferior con borde anterior blanco cremoso, borde inferior pardo oscuro; vientre blanco cremoso; superficie ventral de los miembros traseros crema anaranjado pálido; talón manchado de crema blancuzco. Iris negro; membrana palpebral con los bordes pardo oscuro; resto de la membrana translúcida.
Medidas del holotipo: El onomatóforo es una hembra adulta (huevos ováricos presentes) con las siguientes medidas (en mm): longitud rostro-cloacal 54.8, longitud cabeza 18.0, ancho cabeza 20.1, longitud tibia 27.2, longitud pata 20.1, longitud mano 16.9, longitud horizontal del ojo 5.8, longitud horizontal del tímpano 4.3, distancia ojo-narina 5.4, distancia hocico-ojo 7.4. En el Cuadro I se presentan las medidas de la serie de paratopotipos.
| | | | | | | | | | |
| | | | | | | | | | |
| | | | | | | | | | |
| | | | | | | | | | |
Variación: Todos los ejemplares muestran el labio superior blanco o crema inmaculado. El ejemplar EBRG 3422, una hembra inmadura (con oviductos no convolutos), posee una banda dorsolateral pardo cremosa algo difusa desde el borde posterior del ojo hasta la mitad del cuerpo; lo mismo se observa en el juvenil EBRG 3431, excepto que en éste el color blanco se extiende un poco más allá de la mitad del cuerpo y presenta el vientre de un tono más anaranjado que en los adultos. En los ejemplares EBRG 3426 y 3432 (hembras adultas) también muestran las líneas pálidas indistintas. El ejemplar EBRG 3432, tiene una coloración dorsal gris cremoso pálido con los flancos ligeramente más oscuros que el dorso. El ejemplar EBRG 3426 exhibe unas pocas manchas pálidas dorsalmente. El vientre varía entre crema inmaculado hasta crema anaranjado mate; en algunos ejemplares se transparenta la coloración negra del contenido estomacal o los huevos oscuros contenidos en la cavidad abdominal. La lengua es generalmente redondeada, en un ejemplar es cordiforme (EBRG 3423) y en otros dos (EBRG 3426, 3432) presenta una pequeña escotadura en su extremo posterior. Todos los ejemplares muestran la abertura cloacal cubierta dorsalmente por un pliegue carnoso y dos tubérculos grandes, romos, separados por un profundo surco medio ventral. En unos pocos ejemplares se observaron un par de tubérculos grandes y elevados en el vértice dorso-posterior de la región púbica, algo por encima de la región cloacal. Los tubérculos subarticulares de los dedos de la mano III y IV varían entre sencillos, bífidos o divididos en una o ambas manos a la vez. Todo los ejemplares presentaron una mancha redondeada, blanca, pequeña, en la cara dorso-posterior del muslo, próximo a la articulación tibio-femoral; igualmente, todos los ejemplares mostraron el talón manchado de blanco sucio. El rudimento de polex es visible en machos y hembras sin evidencias de queratinización.
Todos los animales muestran la característica del hocico corto y de forma semicircular en vista dorsal. La distancia ojo-narina es siempre menor que la distancia hocico-ojo, excepto en el ejemplar EBRG 3426, en el cual ambas dimensiones son prácticamente iguales. La relación DON/DHO en siete ejemplares (no se incluyó a EBRG 3426) es 0.85 ± 0.06 (0.75-0.93).
Distribución e historia natural: Actualmente la especie sólo se conoce de la localidad tipo (ver Fig. 4). Huber & Alarcón (1988) clasificaron esta área como "Bosque Nublado Costero", presente desde los 1000 m hacia arriba. Además, según Huber & Alarcón (1988), la Sierra de San Luis está rodeada por amplias zonas de "Arbustales xerófilos espinosos", "Tierras agrícolas" fuertemente perturbadas y por "Bosques tropófilos bajos y medios, deciduos". Estas condiciones sugieren la posibilidad de que T. celsae sea endémica de la Sierra de San Luis.
El holotipo y cuatro paratipos (EBRG 3423-26, 3430, 3432), presentan los oviductos con huevos maduros, bicoloreados pardo oscuro y crema. No se midieron ni se contaron. Una hembra (EBRG 3422) contenía huevos pequeños de color castaño pálido. En tres ejemplares (EBRG 3427-3429), no se detectaron hendiduras vocales ("vocal slits"), pero los testículos son enormes (la longitud de los testículos varió entre 5.8 y 7.5 mm), lo que representa entre el 13.1% y 18.2% de la LRC de los machos. En EBRG 3431, no se observaron las gónadas ni ningún otro caracter sexual secundario, lo que sugiere es un juvenil. No se conoce el renacuajos ni el canto.
A un ejemplar (EBRG 3423) presentó amputación completa del cuarto dedo manual izquierdo; se puede observar, en el lugar de la inserción del dedo, el tejido cicatrizado con los bordes irregulares por lo que, presumimos, este ejemplar sobrevivió a un evento de depredación.
Etimología: El nombre de la especie, "celsae", es un matronímico en reconocimiento a J. Celsa Señaris, quien es coautora de la revisión que culminó en la descripción del género Tepuihyla, por su incansable y destacada labor investigadora de la anurofauna tepuyana y a la incondicional amistad y generosa colaboración que siempre le ha brindado a los autores.
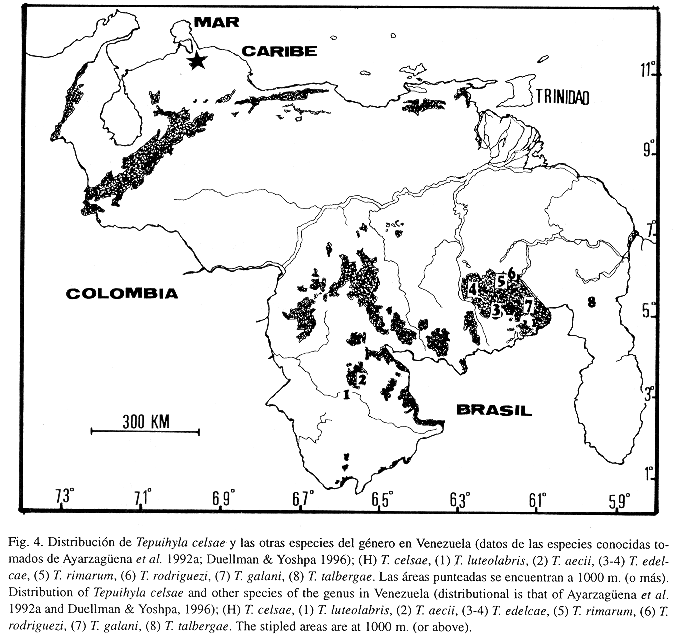
El hecho de que Tepuihyla celsae no haya sido detectada, luego de varios años de muestreo en la zona, sugiere que es una especie escasa, o que prefiere microhábitats específicos o de difícil acceso dentro del bosque nublado o exhiba un comportamiento sumamente discreto (o incluso una combinación de todas las posibilidades mencionadas). Lamentablemente, no se conocen detalles acerca de las condiciones de colecta ni del microhábitat de la especie. Probablemente se trate de una especie endémica a las partes más altas de la Sierra de San Luis. Las siete especies hasta hoy conocidas de Tepuihyla han sido encontradas dentro de plantas de Bromelia (Bromeliaceae) terrestres o en hendiduras en las rocas (Ayarzagüena et al, 1992a; Duellman & Yoshpa 1996).
Las relaciones interespecíficas en Tepuihyla se mantienen indefinidas. Tepuihyla luteolabris y T. celsae comparten la textura tuberculosa de la piel, la ausencia de espículas y la presencia de una notable banda pálida supralabial (esta última característica presente también en T. aecii, y T. talbergae). Tepuihyla luteolabris se conoce actualmente sólo de las partes altas (> 2000 m) del Cerro Marahuaca, en el estado Amazonas, al sur de Venezuela (Ayarzagüena et al. 1992a).
Las relaciones intergenéricas de Tepuihyla, Ayarzagüena et al. (1992b) vinculan estrechamente a ésta con Scinax a partir de la peculiar fórmula de palmeadura manual y pedal, las proporciones del cráneo, la forma de los discos terminales de los dígitos y el esperma con doble filamento; Duellman & Yoshpa (1996) consideraron que la morfología del esperma con doble filamento era la única sinapomorfia que relacionaría a Tepuihyla con Scinax y Sphaenorhynchus y que, la osteología y la presencia de un saco vocal bífido coloca a Tepuihyla en una situación intermedia entre Hyla y Osteocephalus, concluyendo que al menos una de las dos condiciones debe ser producto de convergencia evolutiva.
Una radiografía del holotipo de T. celsae revela que las diapófisis sacras son estrechas e inclinadas hacia atrás, similares a la ilustración de Duellman & Wiens (1992: 11, Fig. 5B) para el género Scinax y considerado como una entre tres sinapomorfias del clado Scinas-Scarthyla-Sphaenorhynchus. Duellman & Yoshpa (1996), aparentemente no tomaron en cuenta los discos terminales dilatados, truncados, más anchos que largos y la ausencia de palmeadura entre los dedos pedales I y II descritos para Tepuihyla, pero que también fueron postulados por Duellman & Wiens (1992), como sinapomorfias (además de la región loreal deprimida) que definen la monofilia de Scinax. A pesar que las relaciones del género Tepuihyla parecen confusas, la información publicada y las pocas evidencias disponibles sugieren una estrecha relación entre Tepuihyla y el clado Scinax-Scarthyla-Sphaenorhynchus y que, en consecuencia, el saco vocal bífido (única semejanza que lo vincularía con Osteocephalus) sería el resultado de convergencia evolutiva.
Biogeografía: A pesar de las distancias relativamente equidistantes, la Sierra de San Luís parece haber tenido una relación biogeográfica más estrecha con la Cordillera de la Costa que con los Andes. Esta relación es notable desde el punto de vista florístico, por el gran número de plantas endémicas compartidas. La segunda relación es la afinidad de plantas con distribuciones disyuntas entre la región guyano-amazónica y el conjunto montañoso del norte de Venezuela conformado por la Sierra de San Luis y la Cordillera de la Costa. Estas afinidades van desde las más débiles en el Oeste, hasta las más pronunciadas en la Península de Paria, al Este, sobre todo en los Cerros El Humo y Patao, que destacan por su estrecha afinidad florística con la región guyano-amazónica (Steyermark 1979).
Las afinidades biogeográficas del conjunto Sierra de San Luís y Cordillera de la Costa pueden clasificarse en un patrón disyunto con dos categorías principales. La primera incluye animales y plantas que no se han diferenciado específica o subespecíficamente, y que presentan una distribución disyunta con poblaciones de la región guyano-amazónica. Ejemplo de ello son, según Steyermark (1979, 1986), las plantas Cespedesia spathulata, Gloespermiun sphaerocarpum, Graffenriela weddellii, Platycentrum clidemioides, Anomospermum reticulata, Macrolobium colombianum, Quiina cruegeriana y Schlegelia parviflora. Sobre la fauna, las distribuciones de aves en Meyer & Phelps (1978) sugieren que los pericos Pyrrhura leucotis y Nannopsittaca panychlora, el saltarín verde (Pipitres chloris) y el atrapamoscas verdoso Xanthiomyias virescens (= Phyllomyias virescens según Lentino 1997) se ajustan a este patrón. Müller (1973) notó que 21 de las 77 aves endémicas de los tepuyes en la Guayana venezolana tenían parientes más cercanos en la porción central de la Cordillera de la Costa, a altitudes sobre 1400-1500. En los mamíferos este mismo patrón disyunto es exhibido por algunos marsupiales (Caluromys philander, Marmosops parvidens y Gracilianus emiliae [Linares 1998, Pérez-Hernández et al. 1994]) y un artiodáctilo (Mazama gouazoubira [Linares 1998]); y entre las serpientes (Roze 1966), por Pseustes sulphureus y Xenodon severus. Gonatodes humeralis es una lagartija que, aunque tiene una distribución predominantemente amazónica (Avila-Pires 1995), extiende su área de distribución hasta la porción central de la Cordillera de la Costa, en los estados Miranda, Aragua y Falcón (Rivero-Blanco 1979, Manzanilla et al. 1996); otro ejemplo de distribución similar lo presenta Plica plica (Manzanilla et al. 1996, A. Mijares-Urrutia datos no publicados). Hyla luteoocellata y Phyllomedusa trinitatis muestran una distribución principal en la Cordillera de la Costa (La Marca 1992, Manzanilla et al. 1995), pero alcanzan la región de Guayana.
La segunda categoría incluye géneros con por lo menos una especie diferenciada en la Cordillera de la Costa y con el resto de taxa relacionados en la región guyano-amazónica. Entre las plantas destacan Hedyosmum gentryi var. A, Rhynchospora nardifolia y Fimbristylis aspera (Robert Wingfield comunicación personal). Entre los anfibios, además de la nueva Tepuihyla descrita aquí, destaca Colostethus sp., mencionado previamente en la literatura como C. brunneus y que se refiere en realidad a una especie de la Amazonía con la que parece estar más estrechamente relacionada filogenéticamente (La Marca 1997b).
Otro caso que pudiera considerarse dentro de esta segunda categoria serían el de especies en el género Flectonotus, cuya distribución abarca la Serranía de Paria (extremo oriental de Venezuela, F. fitzgeraldi) y la Cordillera de la Costa, Sierra de San Luis y los Andes de Venezuela (F. pygmaeus), y que están más estrechamente relacionadas con el género Stefania, reconocido como endémico de las cumbras tepuyanas o bosques bajos adyacentes en Venezuela y Guyana (sensu Duellman & Gray 1983, Duellman & Hoogmoed 1984, Señaris et al. 1997 ["1996"]).
Estas distribuciones y afinidades indican una conexión remota entre las biotas de la Cordillera de la Costa y la del sur de Venezuela. Ésta se estableció presumiblemente (Steyermark 1979) durante una de las fases húmedas del Pleistoceno, a través de "lenguas" y remanentes de selvas de galería, persistentes en sectores no inundados de los Llanos y/o el drenaje Orinoco-Río Negro. Según Steyermark (1979), durante los estadíos interglaciales más cálidos, la biota de tierras bajas emigraría hacia las tierras altas, encontrando ambientes más fríos pero igualmente húmedos en las cumbres con selvas nubladas de la Cordillera de la Costa. La dispersión de elementos de ambientes húmedos entre la región andina y la Cordillera de la Costa y/o la región guyano-amazónica probablemente estuvo retardada o impedida por condiciones áridas y semiáridas de la depresión de Lara-Yaracuy, y de los Llanos, respectivamente, durante las fases más frías y secas del Pleistoceno. No descartamos la posibilidad que la conexión entre la Cordillera de la Costa y el sur de Venezuela se haya dado en forma de "Arco", través de las zonas boscosas del delta del Orinoco.
El registro fósil ofrece indicios de la existencia de una gran y única paleo-provincia ambiental en función del contenido faunístico existente en la zona de Falcón, el cual era muy similar al conocido actualmente en los ríos Orinoco y Amazonas (Audemard 1997).
Agradecemos a Ramón Rivero y Francisco Bisbal por el préstamo de los ejemplares estudiados y su permanente colaboración. A Juan Alfredo Mirt y Johnny Yánez del Centro de Microscopía Electrónica de la Universidad Francisco de Miranda por su valiosa ayuda con las fotografías. Angela Cirilli permitió una radiografía del holotipo. William E. Duellman hizo comentarios valiosos sobre la taxonomía de esta nueva especie. Celsi Señaris confirmó la identidad genérica. La serie tipo fue coleccionada por personal de la Colección de Aves W. H. Phelps, Caracas, y cedida a la Colección de Vertebrados de la Estación Biológica de Rancho Grande; a Ramón A. Rivero (EBRG) por comunicar la existencia de estos ejemplares. Robert Wingfield colaboró con comentarios de especies botánicas de la Sierra de San Luis. Este trabajo es parte del proyecto S1-9112-030, financiado parcialmente por Fundacite-Falcón (para AMU).
Resumen
Se describe una nueva especie de rana arborícola, Tepuihyla celsae (Hylidae), proveniente del bosque nublado (1250 m) de la Sierra de San Luis, Estado Falcón, noroeste de Venezuela. Esta nueva especie se distingue de sus congéneres principalmente por el color dorsal castaño oscuro con unas pocas manchas blancas pequeñas y dispersas, labio superior blanco inmaculado, piel dorsal densamente tuberculosa en ambos sexos, distancia ojo-narina más corta que la longitud horizontal del ojo, dedos de la mano sin membrana, membrana axilar ausente. Esta nueva Tepuihyla representa el registro más septentrional del género, aportando evidencias para establecer una afinidad biogeográfica del conjunto Sierra de San Luís y Cordillera de la Costa con la región guyano-amazónica.
Referencias
Audemard, F. 1997. Importancia geológica del Estado Falcón. Abran Paso (Venezuela) 2: 31-32. [ Links ]
Ávila-Pires, T.C.S. 1995. Lizards of Brazilian Amazonia (Reptilia: Squamata). Zool. Verh. Leiden 299: 1-706. [ Links ]
Ayarzagüena, J., J. C. Señaris & S. Gorzula. 1992a. El grupo Osteocephalus rodriguezi de las tierras altas de la Guayana Venezolana: descripción de cinco especies nuevas. Mem. Soc. Cienc. Nat. La Salle 52: 113-142. [ Links ]
Ayarzagüena, J., J. C. Señaris & S. Gorzula. 1992b. Un nuevo género para las especies del "grupo Osteocephalus rodriguezi" (Anura: Hylidae). Mem. Soc. Cienc. Nat. La Salle 52: 213-221. [ Links ]
Burke, K., C. Cooper, J. F. Dewey, P. Mann & J. L. Pindell. 1984. Caribbean tectonics and relative plate motions. p. 31-64. In W. E. Bonini, R. B. Hargraves & R. Shagam (eds.). The Caribbean-South America Plate Boundary and Regional Tectonics. Memoirs Geol. Soc. Amer., 162. [ Links ]
Díaz de Gamero, L. 1977. Estratigrafía y micropaleontología del Oligoceno y Mioceno inferior del centro de la Cuenca de Falcón, Venezuela. Geos 22: 2-50. [ Links ]
Duellman, W. E. & P. Gray. 1983. Developmental biology and systematics of the egg-brooding hylid frogs, genera Flectonotus and Fritziana. Herpetologica 39: 333-359. [ Links ]
Duellman, W. E. & M. S. Hoogmoed. 1984. The taxonomy and phylogenetic relationships of the hylid frog genus Stefania. Misc. Publ. Univ. Kansas Mus. Nat. Hist. 75: 1-39. [ Links ]
Duellman, W. E. & M. S. Hoogmoed. 1992. Some hylid frogs from the Guiana highlands, northeastern South America: new species, distributional records, and a generic reallocation. Occas. Pap. Mus. Nat. Hist. Univ. Kansas 147: 1-21. [ Links ]
Duellman, W. E. & J. J. Wiens. 1992. The status of the hylid frog genus Ololygon and the recognition of Scinax Wagler, 1830. Occas. Pap. Mus. Nat. Hist. Univ. Kansas 151: 1-23. [ Links ]
Duellman, W. E. & M. Yoshpa. 1996. A new species of Tepuihyla (Anura: Hylidae) from Guyana. Herpetologica 52: 275-281. [ Links ]
González de Juana, C., J. Iturralde & X. Picard. 1980. Geología de Venezuela y de sus cuencas petrolíferas. Edit. Foninves, Caracas. Vol. 1: 407 p. et Vol 2: 624 p. [ Links ]
Heyer, `W. R., A. S. Rand, C. A. G. da Cruz, O. L. Peixoto & C. E. Nelson. 1990. Frogs of Boracéia. Arq. Zool., São Paulo 31: 231-410. [ Links ]
Huber, O. & C. A. Alarcón. 1988. Mapa de Vegetación de Venezuela. Caracas. Escala 1: 2.000.000, 95x70.5 cm. [ Links ]
La Marca, E., 1992. Catálogo taxonómico, biogeográfico y bibliográfico de las ranas de Venezuela. Cuadernos Geográficos (Univ. Los Andes, Mérida) 9: 1-197. [ Links ]
La Marca, E., 1994.- Taxonomy of the frogs of the genus Mannophryne (Amphibia: Anura: Dendrobatidae). Publ. Soc. Amigos de Doñana 4: 1-75. [ Links ]
La Marca, E. 1997a. Origen y Evolución Geológica de la Cordillera de Mérida, Andes de Venezuela. Cuadernos de la Escuela de Geografía (Segunda Etapa, Univ. Los Andes, Mérida) 1: 110. [ Links ]
La Marca, E. 1997b ("1996"). Ranas del género Colostethus (Amphibia : Anura; Dendrobatidae) de la Guayana venezolana, con la descripción de siete especies nuevas. Publ. Asoc. Amigos de Doñana 9: 1-64. [ Links ]
La Marca, E. 1997c. Lista actualizada de los anfibios de Venezuela. p. 103-120. In E. La Marca (ed.). Vertebrados Actuales y Fósiles de Venezuela. Museo de Ciencia y Tecnología, Mérida, Venezuela. [ Links ]
Lentino, M. 1997. Lista actualizada de las aves de Venezuela. p. 145-202. In E. La Marca (ed.). Vertebrados Actuales y Fósiles de Venezuela. Museo de Ciencia y Tecnología, Mérida, Venezuela. [ Links ]
Linares, O. J. 1998. Mamíferos de Venezuela. Sociedad Conservacionista Audubon de Venezuela, Caracas. 691 p. [ Links ]
Mann, P., C. Schubert & K. Burke. 1990. Review of Caribbean neotectonics. p. 307-338. In The Geology of North America. Vol. H. The Caribbean Region. The Geological Society of America. [ Links ]
Manzanilla Puppo, J., A. Fernández Badillo, E. La Marca & R. Visbal García. 1995. Fauna del Parque Nacional Henri Pittier, Venezuela: composición y distribución de los anfibios. Acta Cient. Venez. 46: 1-9. [ Links ]
Manzanilla Puppo, J., A. Fernández Badillo & R. Visbal García. 1996. Fauna del Parque Nacional Henri Pittier, Venezuela: composición y distribución de los reptiles. Acta Cient. Venez. 47: 1-12. [ Links ]
Meyer de Schauense, R. & W. H. Phelps, Jr. 1978. A Guide to the Birds of Venezuela. Princeton Univ.. Princeton, Nueva Jersey. 424 p. [ Links ]
Müller, P. 1973. The dispersal centres of terrestrial vertebrates in the Neotropical Realm. A study in the evolution of the biota and its native landscapes. Biogeographica 2. The Junk, La Haya. [ Links ]
Pérez-Hernández, R., P. Soriano & D. Lew. 1994. Marsupiales de Venezuela. Cuadernos Lagoven, Caracas. 73 p. [ Links ]
Rivero, J. A. 1968. A new species of Hyla (Amphibia, Salientia) from the Venezuelan Guayana. Breviora, Mus. Comp. Zool., Harvard 307: 1-5. [ Links ]
Rivero-Blanco, C. 1979. The neotropical lizards genus Gonatodes (Sauria: Sphaerodactylinae). Tesis Doctoral, Texas A & M University, Station College. 233 p. [ Links ]
Roze, J.A. 1966. La taxonomía y zoogeografía de los ofidios de Venezuela. Ediciones de la Biblioteca, Universidad Central de Venezuela, Caracas. 362 p. [ Links ]
Señaris, J. C., J. Ayarzagüena & S. Gorzula. 1997 ("1996"). Revisión taxonómica del género Stefania (Anura: Hylidae) en Venezuela con la descripción de cinco especies. Publ. Asoc. Amigos de Doñana 7: 1-57. [ Links ]
Steyermark, J. 1979. Plant refuge and dispersal centres in Venezuela : their relict and endemic element. p. 185-221. In K. Larsen & L. B. Holm-Nielsen (eds.). Tropical Botany. Acad.. Londres. [ Links ]
Steyermark, J. 1986. Speciation and endemism in the flora of the Venezuelan tepuis. p. 317-373. In F. Vuilleumier & M. Monasterio (eds.). High Altitude Tropical Biogeography. Oxford Univ., Nueva York. [ Links ]
1 Centro de Investigaciones en Ecología y Zonas Áridas (CIEZA), Universidad Francisco de Miranda,
Apartado 7506, Coro 4101-A, Venezuela. Fax 58-68-524922. Correo-electrónico: amijares@funflc.org.ve
2 Estación Biológica "Dr. Alberto Fernández Yépez", Instituto de Zoología Agrícola, Facultad de Agronomía,
Universidad Central de Venezuela, Apartado 4579, Maracay 2101-A, Venezuela. Fax 58-43-453242.
Correo-electrónico: jmanzanil@yahoo.com
3 Laboratorio de Biogeografía, Facultad de Ciencias Forestales y Ambientales, Universidad de Los
Andes, Apartado 116, Mérida 5101-A, Venezuela. Correo-electrónico: lamarca57@cantv.net

