











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.47 no.4 San José dic. 1999
(Cephalopoda: Loliginidae)
Dwight Arrieche 1
Received. 9- II-1998. Corrected. 07-IV-1999 Accepted. 16-IV-1999
Abstract
I describe the fine structure of the optic gland (OG) in the tropical squid Sepiotheutis sepioidea at different sexual maturity stages. Squids were collected in Mochima National Park, Venezuela (10º 20' 22 " 10º 24' N; 64º 19' 64º 22' W). Morphometric characteristics and the development of reproductive organs at different sexual maturity stages (female n = 101; male n = 105), were significantly different (ANOVA, p < 0.001). The female gonadal index mean (GI = (ovary weight / body weight) x 100) between 0.1 - 2.95 %, and the male GI mean (GI = (testicule weight / body weight) x 100) oscillated between 0.22 - 1.38 %. The OG are a pair of oval organs located at the posterior edge of the optic tract of the cephalopod brain. The OG showed the same cellular types described in other cephalopods, with slight differences. The secretory cells in young individuals present a dendritic profile with thin cytoplasmic processes, numerous circular mitochondriae with tubular crests and abundant ribosomes. At the last stage of maturity the secretory cells contain rough endoplasmic reticulum in the form of dilated cisternae, with an active Golgi apparatus, secreting electron dense granules. Mitochondriae are scarce and lipofuchsin granules show an increase in size. Secretory cell nucleus length and organelle abundance showed oscillations associated with the life cycle.
Key words
Optic gland, cephalopod, Sepiotheutis sepioidea, ultrastructure.
In cephalopods the optic glands (OG) form a pair of oval organs located at the posterior edge of the optic brain tract, above the hilum of the optic lobes (Boycott & Young 1956). OG histology has been described for Loligo opalescens, Octopus vulgaris, O. bimaculatus, Eledone moschata and Sepia officinalis. The OG consists of main or secretory cells, blood vessels, nerve cells and support cells (Björkman 1963, Nishioka et al. 1966, Barber 1967, Bonichon 1967, Froesch 1974 and Mangold & Froesch 1977).
The role of the OG in the cephalopod life cycle is analogous to that of the hypophysis which controls sexual development in vertebrates (Wells & Wells, 1959, 1969, Wells 1960 and Wells et al. 1975). These glands also regulate protein synthesis in the ovaries, stimulate gonadal mitosis, mating behavior, feeding and longevity (Durchon & Richard 1967, O´Dor & Wells 1973, Wells et al. 1975, Wodinsky 1977 and Mangold 1987). The OG hormone is species specific, not sex specific (Wells & Wells 1975), and glandular secretions have periodic oscillations associated with temperature and photoperiod (Defretin & Richard 1967 and Richard 1966, 1967 a, b, 1970). The glands are controlled by a reactive peptidergic inervation (Le Gall et al. 1988), and their activity varies during the life cycle, associated with the multiplication of reproductive cells (Koueta et al. 1995).
The fine structure of the gland suggests that it has several functions. Björkman (1963) suggested that the secretory cells had a high rate of synthesis. The presence of abundant mitochondriae implies that the gland may secrete a steroid hormone (Froesch 1979). In S. officinalis, however a peptidic like hormone was shown to be present in the OG and hemolymph (Koueta et al. 1992). The lipofuchsin granules have been related to the resorption of metabolic residues inside the secretory cells (Froesch & Mangold 1976, Mangold & Froesch 1977 and Froesch et al. 1987).
In this study the fine ultrastructure of the tropical squid Sepiotheutis sepioidea (Blainville, 1823) (Cephalopoda: Loliginidae) was examined in both sexes at different sexual maturity stages, using Transmission Electron Microscope (TEM) in order to compare secretory cell changes during the life cycle of this specie.
Materials and Methods
206 individuals of S. sepioidea were caught (net mesh size 20 mm Ø) in Mochima National Park, Venezuela (10º 20' 22 " 10º 24' N; 64º 19' 64º 22' W). They were acclimatized in tanks (1 500 L) with running sea water, and fed ad lib. for at least a week (Estación José G. Hernández, IDEAFundaciencias). Each specimen was weighed (body weight BW ± 0.05 g) and the dorsal mantle length (DML ± 0.1 cm) was measured before decapitation, followed by dissection of the OG, with the aid of a stereomicroscope. The OG was preserved by immersion as quoted below for light microscopy (LM) and TEM.
The mantle with the reproductive organs was preserved with 4% formalin (24 h), and then washed in running tap water and the reproductive organs were removed and measured (± 0.05 cm; ± 0.0001 g). The sexual maturity stage was determined for each specimen as follows: juvenile (I), immature (II), in maturation (III), mature (IV), and post-reproductive (V). This last was identified only in copulating males, and post-reproductive females were not captured. The sexual maturity stages were determined using the measurements of reproductive organs quoted for this species (Robaina & Voglar 1986, Voglar & Robaina 1987), and the female sexual maturity stages were estimated using the nidamental gland index (NGI = (Nidamental Gland length/ Dorsal Mantle length)) (Durward et al. 1979). Gonadal index (GI) was estimated as the relationship between gonad weight and body weight, in the females the gonadal index was calculated using the formula (GI = (ovary weight / body weight) x 100), and in the males this was calculated as (GI = (testicule weight / body weight) x 100). An analysis of variance (ANOVA) was used to test for significant differences in the biometric parameters between the sexual maturity stages (females) and the dorsal mantle length intervals (males), these data were compared using an a posteriori Scheffe test (Sokal & Rohlf 1981).
For LM, samples including the optic lobe and the optic stalk were immersed in Gilson fixative (2 - 6 h), and then well washed in running tap water, dehydrated and embedded in paraffin. Paraffin sections (6 µm) were stained with Hematoxilyn-Eosin (HE), Periodic acid Schift reaction (PAS) and Herlant tetrachromic stain (Humason 1979). LM sections were analyzed using an image analysis system (Javelin Smart Cam) adapted to a light microscope, with a digital micrometer.
For TEM, the brain was prefixed by immersion in either of the following fixatives (30 min.) with some drops of 0.1% (w/v) methylene blue (vital stain)(Bern, H., pers. com.): 2.5% glutaraldehyde (GA) in 0.1M phosphate buffer saline pH 7.8; 2.5 % GA in 0.1M sodium cacodylate buffer pH 7.8; 3% GA, 20% sucrose (w/v), 0.5% CaCl2, 1% K2Cr2O7, 0.1% picric acid in 0.1 M sodium cacodylate buffer (Evans at al. 1976). The brain was then washed in the same buffer, and the OG dissected from the cerebral tissue followed by one hour fixation in fresh fixative. The OG was washed, post-fixed (1% OsO4, 90 min.), in bloc stained (2% uranyl acetate, 30 min.), and processed for conventional TEM. Thin sections were contrasted with uranyl acetate and lead citrate (Reynolds 1963), and observed in a H-600 TEM.
Results
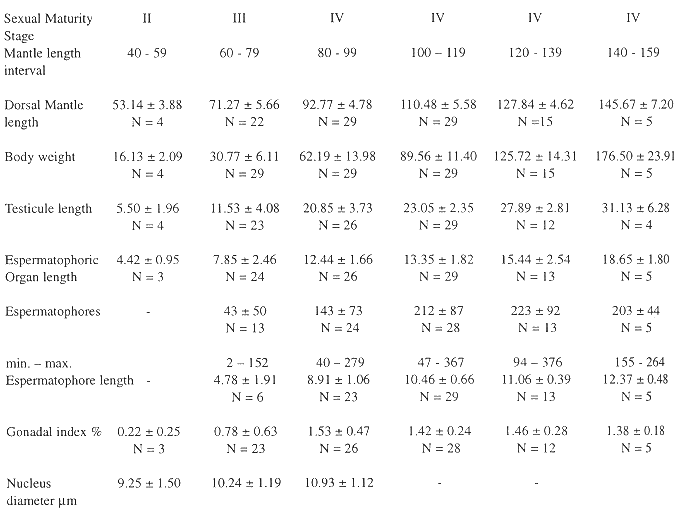
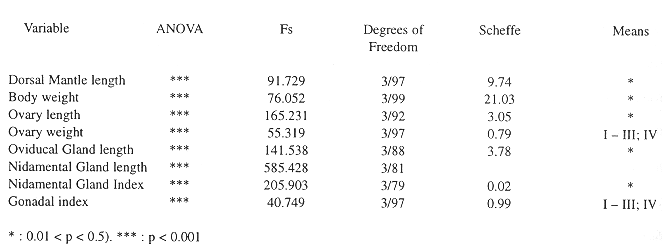
The OG is located over the optic stalk close to the optic lobe, it is covered by a fine connective layer, and has an elongate shape. The OG histology is well differentiated from the cerebral tissue, the OG cells are in a compact mass, whereas cerebral cells are larger, and form layers surrounding eosinophylic nerve fibers (Fig. 1). At the juvenile maturity stage the OG is almost undifferentiated from the cerebral tissue under a stereoscope, having a reddish coloration in individuals with a larger dorsal mantle length, mainly mature females. The female gonadal index (GI) oscillated between 0.1 - 2.95 %, and the nidamental gland index between 0.9 - 0.28 %, with significant differences between the sexual maturity stages (Table 1). The reproductive organs in the males showed a tendency to increase with dorsal mantle length, but the spermatophore number was associated with reproductive behaviour, GI oscillated between 0.22 - 1.38 % (Table 2). These data had significant differences (ANOVA, p < 0.001), but in an a posteriori Scheffe test the means did not differ (Table 3, 4).
Biometric parameters of the female reproductive organs in the different sexual maturity stages (average ± standard deviation; N = sample size). The reproductive organs show a tendency for linear increase in size between the sexual maturity stages, including the diameter of the nucleus. Sample size is indicated below the mean and standard deviation.

Biometric parameters of the male reproductive organs in the different sexual maturity stages. The individuals were grouped in mantle length intervals, the nucleus diameter was averaged at the fourth maturity stage. The oscillation of the espermatophores at the fourth maturity stage is associated with copulation. Sample size is indicated below the mean and standard deviation.
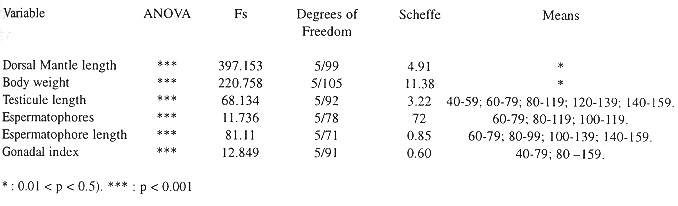
Analysis of variance (ANOVA) and a posteriori Scheffe test of female morphometric parameters at the different sexual
maturity stages.
Analysis of variance (ANOVA) and a posteriori Scheffe test of the males morphometric parameters at the different sexual maturity stages.
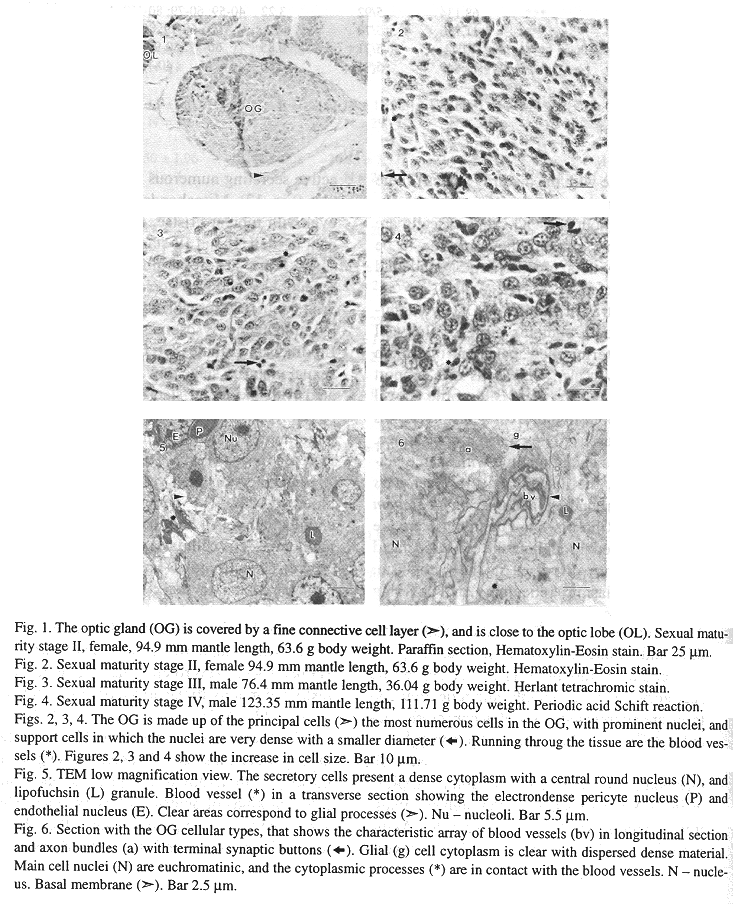
Paraffin sections give a clear picture of OG organization. The secretory cells are the most numerous cells, they are ovoid with cytoplasmic processes (18.5 25.6 *m), and the nucleus (9.11 11.38 *m) is circular with a dense nucleoli, both showing a tendency to increase in size with the maturity stages (Table 1, 2). The blood vessels are prominently dispersed throughout the OG. Support cells are scarce, in some sections they are seen to be aggregated, with a characteristic round and basophilic nucleus (Figs. 1, 2, 3, 4).
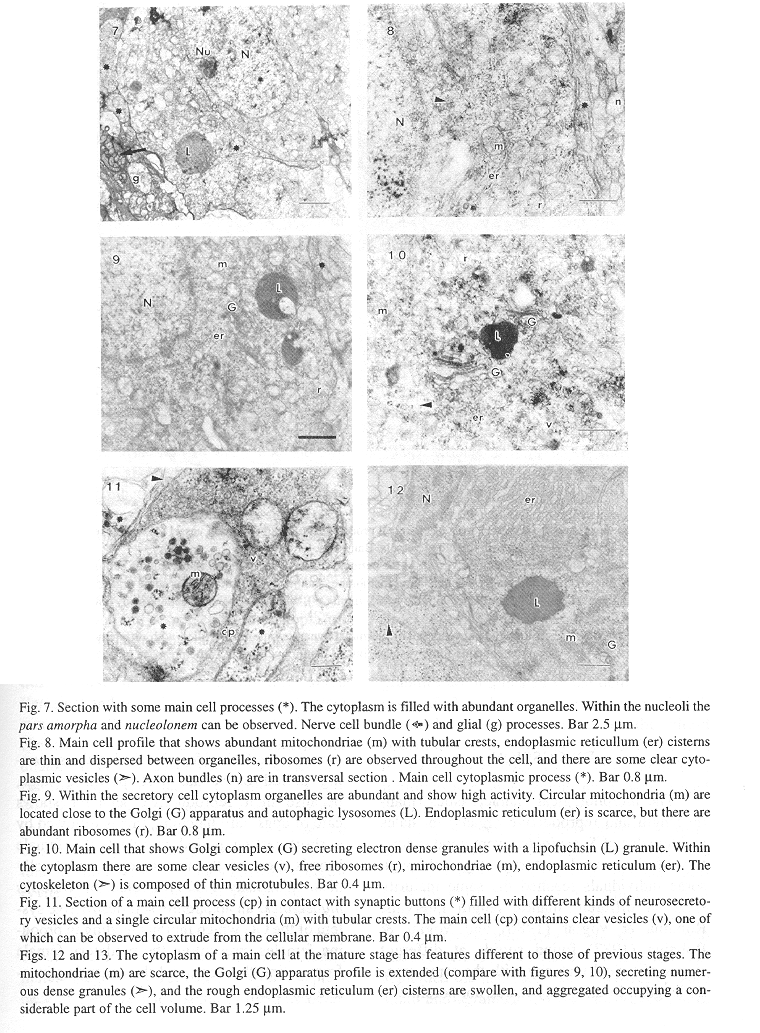
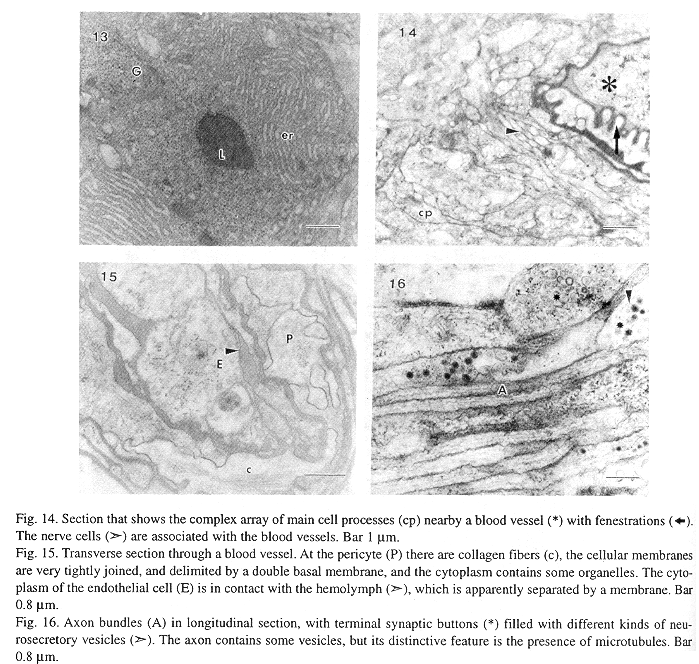
At the ultrastructural level OG cytoarchitecture denotes a complex array of fine cell processes intermingled between the main cells, blood vessels, nerve cells and support cells (Figs. 5, 6). The main cell profile is oval with thin cytoplasmic processes, its organelles are very numerous giving a dense appearance. The nucleus is euchromatinic with scarce heterochromatin, the nucleoli have a clear pars amorpha surrounding a darker nucleolonem (Figs. 5, 7). Ribosomes are numerous, and are found dispersed throughout the cell, associated with polysomes dispersed in the cytoplasm, rough endoplasmic reticulum, and covering clear cytoplasmic vesicles (Figs. 8, 10, 11). The rough endoplasmic reticulum cisterns are thin, and extend from the nuclear envelope to the cellular membrane (Figs. 8, 9, 19), at the secretory stage the endoplasmic reticulum cisterns are aggregated in swollen concentric saccules (Figs. 12, 13). The Golgi apparatus is active, it secretes clear and electrondense vesicles and there are at least two Golgi complexes in each cell associated with rough endoplasmic reticulum, and lipofuchsin granules. At the mature sexual stage (IV) the Golgi is very active secreting numerous electrondense granules (Fig. 12). Mitochondriae are elongated or oval with tubular cristae, they are very abundant in the I, II and III stages, whilst in the IV stage there are few mitochondriae. These reductions could be associated with mitochondrial matrix desorganization and destruction by autophagic lysosomes (Figs. 8, 9, 12). Lipofuchsin granules are found from the third sexual maturity stage onwards, and are associated with the Golgi stacks, these profiles are electron dense with clear inclusions, and in the larger mantle length specimens they shown an increase in size (Figs. 7, 9, 12).
This gland is well vascularized with large blood vessels. The pericyte cells cover the endothelial cells, and have a double basal membrane. The cellular membranes are tightly joined in an irregular fashion, and there are collagen fibers in the cytoplasm. The cytoplasm of the endothelial cells is not easily identified because the lumen has an electrondense hemolymph which occupies nearly the entire space. Nevertheles the organelles are restricted mainly to the area close to the nucleus (Fig. 13, 14, 15).
The nerve cells are distributed between the principal cells and blood vessels in long axon bundles with terminal synaptic buttons with different kinds of neurosecretory vesicles. The axon bundles are tightly packed, they have long microtubules, and some microvesicles. The synaptic buttons are in contact with axon bundles, secretory cells and blood vessels, and there were no synaptic electron dense contacts identified. The neurosecretory vesicles were classified according to electron density and size in: small agranular (35 - 70 nm), agranular (90 - 150 nm), moderate to very dense (95 - 130 nm) and dense with a smooth limiting membrane (90 - 170 nm) (Figs. 6, 11, 14, 16). Glial cells are intermingled between the OG cells, with long cytoplasmic processes, their cytoplasm is clear with a particulate material and few organelles, some mitochondriae, vacuoles and vesicles (Figs. 6, 11, 16).
Discussion
The morphological characteristics (mantle length and reproductive organs) in males and females of S. sepioidea were good indicators of the sexual maturity stage, although some individuals reached the same maturity stage at different mantle length intervals. Robaina & Voglar (1986) and Voglar & Robaina (1987) found squids at different stages of sexual maturity with the same mantle lengths. In this study it was not possible to examine the OG of females during the spawning period or at the post-spawning stage, because their reproductive behavior shows care of eggs until hatching, this is followed by a short resting period in which the female broods most of the mature eggs and finally dies. Wodinsky (1977) quoted in Octopus humeelincki an apparently self destructive system of the OG which inhibits feeding in brooding females, preventing the predatory female from eating the eggs, and thus ensuring juvenile survival.
The OG shows a highly vascularized tissue and fine structure sections from S. Sepioidea were improved by modifying some combinations of fixatives and schedules. The fixer quoted by Evans et al. (1976) followed by extensive washings gave the best tissue preservation, while the aldehyde prepared in phosphate buffer saline with filtered sea water resulted in a bad preservation. Problems were also reported with O. vulgaris and S. officinalis OG fine structure by Defretin & Richard (1967) and Nishioka et al. (1970). In S. Sepioidea a hematonervous barrier built by the cellular wall, and gap junctions (zonula ocludens) between the endothelial cell and the perycite may halt the difussion of the fixative in the squid giant axon (Villegas & Villegas 1968). Villegas (pers. com.) obtained the best preservation by immersion instead of perfusion, this latter being the suggested procedure for delicate tissues.
The fine structure of the tropical squid S. sepioidea OG, showed the same cellular types described in other cephalopods, with some distinctive features. S. sepioidea secretory cells showed differences in organelle abundance between the sexual maturity stages: at the juvenile and immature stages there are numerous mitochondriae, endoplasmic reticulum cisterns are thin and dispersed in the cytoplasm, the Golgi apparatus has a curved profile and actively secretes granules of different densities, whilst in the IV maturation stage (females) and reproductive individuals (males) secretory cells are almost depleted of mitochondriae, endoplasmic reticulum cisterns are dilated and well developed in a concentric fashion, and the Golgi complex actively secretes electron dense granules which are dispersed in the cytoplasm. Some of these are close to the lipofuchsin granules and to the cellular membrane, suggesting a possible secretory mechanism through the blood vessels.
S. sepioidea OG secretory activity was demonstrated by the production of electron dense secretory granules in the sexually mature stage (Figs. 12, 13). However, the presence of clear vesicles, some of them associated with polysomes, could indicate the existence of other secretory processes in the life cycle, as seen by the extrusion of a clear vesicle close to a synaptic button (Fig. 11). Moreover, the cellular characteristics indicate active cells, with euchromatinic nuclei, an abundance of organelles, and an increase in lipofuchsin volume and cell size with dorsal mantle length.
Defretin & Richard (1967) demonstrated that when S. officinalis is kept in a dark photoperiod the OG secretory cells produce three kinds of secretory granules, suggesting that they are filled with the same product in different stages of synthesis. Octopus vulgaris OG enlargment occurred both in the nucleus and secretory cell cytoplasm after cerebral surgery (Wells 1964). Eledone cirrhosa OG enlargement was statisticaly associated with female gonad maturation, and it is suspected that this process occurs normally (Boyle & Thorpe 1984). Koueta et al.(1995) described cellular changes in S. officinalis OG histology during sexual maturation; and found that main cell secretory activity was correlated with a peak of mitogenic activity in both sexes.
Lipofuchsin granules in the OG of some octopods are numerous, and measured up to several micrometers in diameter. Mangold & Froesch (1977) did not find a correlation between the size of lipofuchsin granules and sexual development in females. Lipofuchsin granules are implicated in the defense mechanism of O. vulgaris in the presence of high molecular weight molecules in the blood stream (Froesch & Mangold 1976, Froesch et al. 1978 and Froesch 1979). In S. sepiodea a lipofuchsin granule engulfing a degenerated mitochondriae was observed.
The presence of different types of neurosecretory microvesicles in the axon bundles and synaptic buttons in the OG of S. sepioidea indicates the existence of different stimuli controlling OG activity. These stimuli were reported as excitatory or inhibitory (Nishioka et al. 1966 and Froesch 1974). In the stellate ganglion of S. sepioidea the neurosecretory microvesicles are small and agranular (50 - 80 nm) (Castejón & Villegas 1964), in the stellate ganglion of Loligo vulgaris there are both small agranular vesicles (40 - 60 nm) and large agranular vesicles (average 65 nm) (Gervasio et al. 1971). In the OG of Loligo opalescens the more common granular vesicles have a loose limiting membrane (80 - 160 nm), and a few axon-like processes containing dense vesicles (100 - 300 nm) with a smooth limiting membrane (Nishioka et al. 1970).
Field populations of S. sepioidea have a steady increase of the OG secretory cell diameter in both sexes. Sexual maturation in octopus was associated with a gross enlargment of the OG and a change in colour from cream to bright orange after optic nerve surgery (Wells & Wells 1959). In S. officinalis growth and sexual development was stimuled by a dark photoperiod and high temperature (Richard 1966, 1967 a, b, Defretin & Richard 1967). Likewise O. vulgaris mature individuals showed more intense histological reactions, and an increase in size in cytoplasm, nucleus and granules in the principal cells (Bonichon 1967). Eledone cirrhosa OG enlargment was statistically associated with the weight of the ovary and oviducal glands (Boyle & Thorpe 1984). Lipofuchsin granules and octopus body weight were well correlated, but the secretory mechanism was not demonstrated (Mangold & Froesch 1977).
Acknowledgements
I thank "Estación Experimental José Gregorio Hernández, Fundaciencias - IDEA", Jesús Rodríguez, Susan Tai, Gilma Hernández de A., Frances Osborn and Osmar Nusetti.
Resumen
Se examinó la ultraestructura fina de la glándula óptica (GO) en varios estadios de desarrollo sexual del calamar tropical Sepiotheutis sepioidea, proveniente de las costas de la Bahía de Mochima, Venezuela (10º 20' 22 " 10º 24' N; 64º 19' 64º 22' W). Los parámetros morfométricos y de los órganos reproductivos (hembras n = 101; machos n = 105), presentaron diferencias significativas entre los estados de maduración sexual (ANOVA, p < 0.001), el índice gonadal promedio en las hembras osciló entre 0.1 - 2.95 %, y en los machos entre 0.22 - 1.38 %. La GO son un par de órganos ovales ubicados en el borde posterior del tracto óptico en el cerebro de los cefalópodos, ésta posee los mismos tipos de células descritos en otros cefalópodos con algunas leves diferencias. La célula principal en el estadio de maduración juvenil posee un perfil dendrítico con delgados procesos del citoplasma, abundantes ribosomas, mitocondrias circulares con crestas tubulares, y un núcleo prominente eucromatínico; en el estadio de adulto las células principales presentan las cisternas del retículo endoplasmático rugoso dilatadas concéntricamente en la célula, el aparato de Golgi es activo con abundantes gránulos electron densos de secreción, las mitocondrias son escasas, y el gránulo de lipofucsina es de mayor tamaño. La dimensión del núcleo de la célula principal y la abundancia de organelos, presentaron oscilaciones en las etapas del ciclo de vida.
References
Barber, V. 1967. A neurosecretory tissue in Octopus. Nature 213:1042-1043. [ Links ]
Björkman, N. 1963. On the ultrastructure of the optic gland in Octopus. J. Ultrastruct. Res. 8:195. [ Links ]
Bonichon, A. 1967. Contribution à l´etude de la neurosécretion et de l´endocrinologie chez les Céphalpodes. I. Octopus vulgaris. Vie Milieu 18:27-263. [ Links ]
Boycott, B. B. & J. Z. Young. 1956. The subpedunculate body and nerve and other organs associated with the optic tracts of cephalopods. Bertil Hanström Zoological Papers 76-105 p. [ Links ]
Boyle, P. R. & R. Thorpe. 1984. Optic gland enlargement and female gonad maturation in a population of the octopus Eledone cirrhosa: a multivariate analysis. Mar. Biol. 79:127-132. [ Links ]
Castejón, O. & G. M. Villegas. 1964, Fine structure of the synaptic contacts in the stellate ganglion of the squid. J. Ultrastruct. Res. 10:585-598. [ Links ]
Defretin, R. & A. Richard 1967. Ultrastructure de la glande optique de Sepia officinalis L. (Mollusques; Céphalopode). Mise en evidence de la sécrétion et de son controle photopériodique. C. R. Acad. Sci., Paris. D. 265:1415-1418. [ Links ]
Durchon, M. & A. Richard. 1967. Étude en culture organotypique du rôle endocrine de la glandule optique dans la maturation ovarienne chez Sepia officinalis L. (Mollusque: Céphalopode). C. R. Acad. Sci., Paris. D. 264:1497-1500. [ Links ]
Durward, R., T. Amaratunga & R. O´Dor. 1979. Maturation index and fecundity for the female squid Illex illecebrosus. ICNAF Res. Bull. 14:67-72. [ Links ]
Evans, P., E. Kravitz & B. Talamo. 1976. Octopamine release at two points along lobster nerve trunks. J. Physiol. 262:71-89. [ Links ]
Froesch, D. 1974. The subpedunculate lobe of the octopus brain: evidence for dual function. Brain Res. 75:277-285. [ Links ]
Froesch, D. 1979. Antigen-induced secretion in the optic gland of Octopus vulgaris. Proc. R. Soc. Lond. B. 205:379-384. [ Links ]
Froesch, D. & K. Mangold. 1976. Uptake of ferritin by the cephalopod optic gland. Cell Tiss. Res. 170:549-551. [ Links ]
Froesch, D., K. Mangold & W. Fritz. 1978. On the turnover of exogenous ferritin in the cephalopod optic gland. A microprobe. Experientia 34:116-117. [ Links ]
Gervasio, A., R. Martin & A. Miralto. 1971. Fine structure of synaptic contacts in the first order giant fiber system of the squid. Z. Zellforsch. 112:85-96. [ Links ]
Humason, G. 1979. Animal Tissue Techniques. W. H. Freeman, San Francisco, California p. 456-461. [ Links ]
Koueta, N., E. Boucaud-Camou & A. M. Renou. 1995. Gonadotropic mitogenic activity of the optic gland of the cuttlefish Sepia officinalis, during sexual maturation. J. Mar. Biol. Ass. U. K. 75:391-404. [ Links ]
Koueta, N., I. Robbins & E. Boucaud-Camou. 1992. Partial characterization of a gonadotropic, mitogenic factor from the optic gland and hemolymph of the cuttlefish Sepia officinalis. L. Comp. Biochem. Physiol. 102A:224-234. [ Links ]
Le Gall, S., C. Feral, J. VanMinnen & C. Marchand. 1988. Evidence for peptidergic innervation of the endocrine optic gland in Sepia by neurons showing FMRF amide like immunoreactivity. Brain Res. 462:82-88. [ Links ]
Mangold, K. 1987. Reproduction. Chapter X. In "Cephalopod Life Cycles". Academic, New York. p. 157-200. [ Links ]
Mangold, K. & D. Froesch. 1977. A reconsideration of factors associated with sexual maturation. Symp. Zool. Soc. Lond. 38:541-555. [ Links ]
Nishioka, R., H. Bern & D. Golding. 1970. Innervation of the cephalopod optic gland. In "Aspects of neuroendocrinology". W. Bargman & B. Schaer (eds). Springer, Berlin. p. 47-54. [ Links ]
Nishioka, R. S., I Yasumasu, A. Packard, H. Bern & J. Z. Young. 1966. Nature of vesicles associated with the nervous system of cephalopods. Z. Zellforsch. 75:301-316. [ Links ]
O´Dor, R. & M. J. Wells. 1973. Yolk protein synthesis in the ovary of Octopus vulgaris and its control by the optic gland gonadotropin. J. Exp. Biol 59:665-674. [ Links ]
Reynolds, 1963. The use of lead citrate of high pH as an electron opaque stain in electron microscopy. J. Cell Biol. 17:208-212. [ Links ]
Richard, A. 1966. La température, facteur essentiel de croissance pour le Céphalopode Sepia officinalis. C. R. Acad. Sci., Paris. D. 263:1138-1141. [ Links ]
Richard, A. 1967a. Action de la température sur l´evolution génitale de Sepia officinalis. C. R. Acad. Sci., Paris. D 263:1998-2001. [ Links ]
Richard, A. 1967b. Rôle de la photopériode dans le determinisme de la maturation génitale femelle de Céphalopode Sepia officinalis L. C. R. Acad. Sci., Paris. D 265:1315-1318.. [ Links ]
Richard, A. 1970. Differentiation sexuelle de Céphalopodes en culture in vitro. Ann. Biol. IX(7, 9-10):409-415. [ Links ]
Robaina, G. & J. F. Voglar. 1986. Maduración sexual de las hembras de Sepiotheutis sepioidea (Blainville, 1823) (Cephalopoda: Loliginidae) en las costas Venezolanas. Inv. Pesq. 50:57-68. [ Links ]
Sokal, R. & F. Rohlf. 1981. Biometry. W. H. Freeman, San Francisco, California, 859 p. [ Links ]
Voglar, J. F. & G. Robaina. 1987. Maduración sexual de los machos del calamar Sepiotheutis sepioidea (Blainville, 1823) en las costas del nororiente Venezolano. Contr. Cient. Ctro. de Invest. Cient. UDO 11:40 pp. [ Links ]
Wells, M. J. 1960. Optic glands implants and the ovary of Octopus. Symp. Zool. Soc. Lond. 2:87-107. [ Links ]
Wells, M., R. O´Dor & S. Buckley. 1975. An in vitro bioassay for a molluscan gonadotropin. J. Exp. Biol. 62:433-436. [ Links ]
Wells, M. & J. Wells. 1959. Hormonal control of sexual maturity in Octopus. J. Exp. Biol. 36:1-33. [ Links ]
Wells, M. & J. Wells. 1969. Pituitary analogue in the Octopus. Nature 222:293-294. [ Links ]
Wells, M. & J. Wells. 1975. Optic glands implants and their effects on the gonads of Octopus. J. Exp. Biol. 62:579-588. [ Links ]
Wodinsky, J. 1977. Hormonal inhibition of feeding in Octopus control by optic gland secretion. Science 198:948-951. [ Links ]
1 Instituto de Investigaciones en Biomedicina y Ciencias Aplicadas, Universidad de Oriente. Cumaná, Sucre, Venezuela. Fax 58-93-514023. Tel. 58-93-514185. E-mail: darriech@cumana.sucre.udo.edu.ve

