










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.47 n.3 San José Sep. 1999
Abstract
During the three months of field work between October and December 1997 by the junior author in the in the Réserve Spéciale dIvohibe and in the Forêt dAnkazomivady-Ambositra, Province of Fianarantsoa, a collection of scorpions was made, mainly with the use of pitfall buckets. The species collected were Grosphus limbatus (Pocock), Grosphus madagascariensis (Gervais), and new species of Tityobuthus Pocock (Scorpiones, Buthidae), which is described from Ivohibe based on two female specimens. Tityobuthus ivohibe n. sp., is the eleventh species described in the genus from Madagascar. Some comments concerning the taxonomic position of the new species are also presented.
Key words
Scorpiones, Buthidae, Tityobuthus, Grosphus, Ivohibe, Madagascar, New species.
Introduction
During three months of field work done between October and December 1997 by the junior author in the Réserve Spéciale (RS) dIvohibe and in the Forêt dAnkazomivady-Ambositra, both in the Province de Fianarantsoa, a reasonable number of scorpions were collected, mainly with the use of pitfall trap lines. Part of this material was studied by the senior author while on a short visit to the Field Museum and the rest was brought to the Paris Museum for further analysis.
These collections contain three species of scorpions, all belonging to the family Buthidae. Two represent already known species in the genus Grosphus Simon 1880, and the third one has been identified as a new species in the genus Tityobuthus Pocock 1893. With the description of this new species, the total number in the genus Tityobuthus is raised to eleven. The increasing number of new species within Tityobuthus suggests that this genus, which can be included among the micro-scorpion genera, may be very rich in species. A similar phenomenon has already been observed in other micro-scorpion genera such as Ananteris Thorell 1891 of the neotropics, in which the number of described species increased from 3 to 19 in the space of twelve years (Lourenço 1993, 1994). In fact, since the publication of the Scorpion Fauna of Madagascar (Lourenço 1996), other new species of Tityobuthus, Microcharmus Lourenço 1995 and Pseudouroplectes Lourenço 1995 have been discovered and described (Lourenço 1997, 1998; Lourenço & Goodman in press). This situation suggests that the scorpion fauna of Madagascar is still far from being well known and that many new taxa remain to be discovered.
Methods
Madagascar, Province de Fianarantsoa, 28 km SSW Ambositra, 4.7 km SW Ambalamanakana, Forêt dAnkazomivady, 20 46.5 S ; 47 10,1 E (1700 m), 6-15/I/1998 (S.M. Goodman). 3 adult females ; one of the females with a brood composed of 8 males and 6 females.
The majority of specimens were collected on hills exposed rock and were found below rock. These hillrocks were out of the forested zone and were generally surrounded by open grasslands composed of introduced species of Graminaceae. This site of collection probably corresponds to the highest one known for scorpions in Madagascar.
Madagascar, Province de Fianarantsoa, exterior northern limit of Réserve Spéciale dIvohibe, along the Hefitany River, 7,5 km ENE Ivohibe, 22 28,2 S - 46 57,6 E (900 m), camp 1, 6-13/X/1997 (S. M. Goodman, coll.). 6 adult males and 1 juvenile female.
As for Grosphus limbatus, specimens were found under rocks.
Holotype female : Madagascar, Province de Fianarantsoa, exterior northern limit of Réserve Spéciale dIvohibe, along Hefitany river, 7,5 km ENE Ivohibe, 22 28,2 S - 46 57,6 E (900 m), camp 1, 6-13/X/1997 (S. Goodman, coll.).
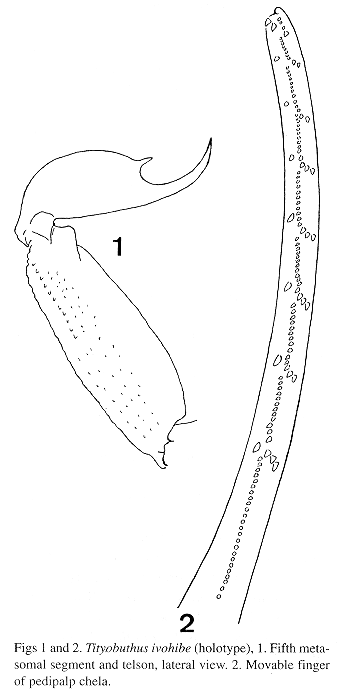
Deposited in the Field Museum of Natural History, Chicago, USA.
Etymology: The specific name is a name placed in apposition to the generic name and makes reference to the type-locality.
Description based on female holotype. Measurements presented in Table I.
Coloration. Basically yellowish, symmetrically marbled with dark brown, producing an overall spotted appearence. Prosoma: carapace yellowish, heavily spotted; eyes surrounded by black pigment. Mesosoma: yellowish with four longitudinal brown stripes. Metasoma: segments I to V yellowish; vesicle yellowish but lighter than segment V. Venter yellow with several spots on all sternites. Sternite III with a very small bright white zone in the posterior region. Chelicerae reddish-yellow with dark spots on the median zone; fingers reddish. Pedipalps yellowish with several spots on femur and tibia; chelae less densely spotted; fingers darker, reddish. Legs yellowish with diffuse fuscous spots.
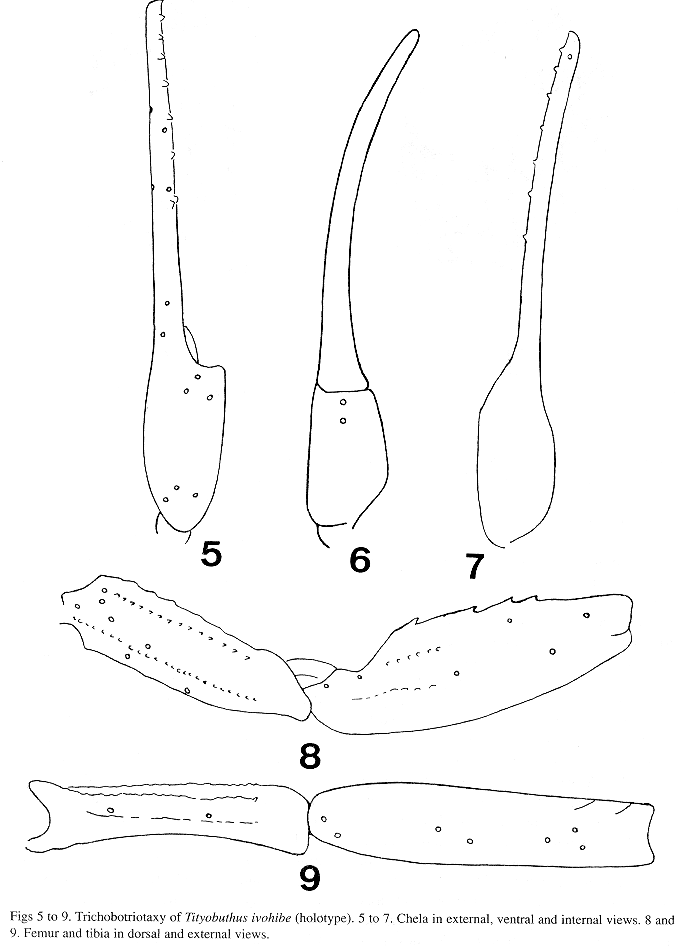
Morphology. Carapace moderately to feebly granular; anterior margin with a median concavity. Anterior median superciliary and posterior median keels moderate to feeble. All furrows moderate to feeble. Median ocular tubercle distinctly anterior to the center; median eyes separated by one ocular diameter. Three pairs of lateral eyes. Sternum subtriangular to pentagonal. Mesosoma: tergites feebly granular. Median keel moderate to feeble in all tergites. Tergite VII pentacarinate. Venter: genital operculum divided longitudinally. Pectines: pectinal tooth count 11-11; basal middle lamellae of the pectines not dilated; fulcra present. Sternites smooth with moderately elongate stigmata; VII without keels. Metasoma: segments I to III with 10 keels, crenulate. Lateral inframedian keels on segments I and II complete, crenulate; incomplet on III. Intercarinal spaces moderate to feebly granular. Segment V feebly granular, almost smooth and rounded. Telson smooth with a moderately long and moderately curved aculeus; subaculear tooth strong and spinoid. Cheliceral dentition characteristic of the genus Tityobuthus (Vachon 1963); ventral aspect of both finger and manus with dense, long setae. Pedipalps: femur pentacarinate; tibia and chelae with some keels but moderately crenulate; internal face of tibia with four spinoid granules; all faces moderate to feebly granular. Movable fingers with 9 oblique rows of granules; internal and external accessory granules present. Trichobothriotaxy; orthobothriotaxy A- (Vachon 1973, 1975). Legs: tarsus with very numerous median fine setae ventrally. Tibial spurs present.
Paratype female: Similar to the holotype female with respect to both coloration and morphology. Pectinal tooth count 11-11. Data for female paratype, same as for the holotype. Deposited in the Muséum National dHistoire Naturelle, Paris.
Taxonomic position of the new species
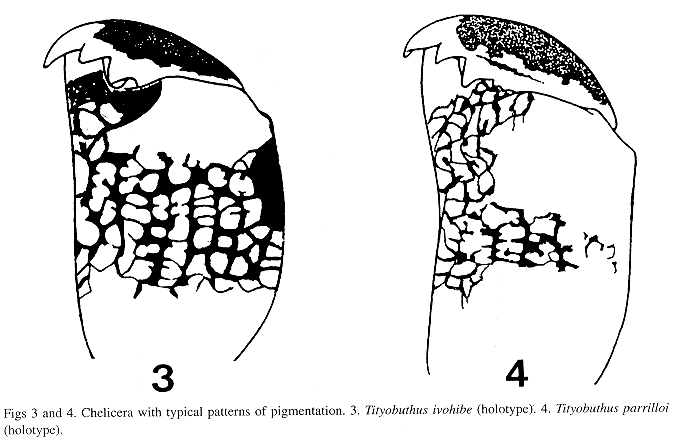
According to the key presented by Lourenço (1996) for the various species in the genus Tityobuthus, the new species T. ivohibe appears to be most closed related to T. parrilloi Lourenço 1996. This latter species is only known from the lowland humid forests of the RNI dAndohahela in the extreme Southeastern portion of the country. The two species can, however, be readily distinguished by the following features: (i) a different pattern of pigmentation of the chelicera (see Figs 3 and 4); (ii) a different pectinal tooth count: 13/14 in T. parrilloi, and 11 in T. ivohibe; (iii) metasomal segments III of T. parrilloi with 8 keels, whereas in the new species segment III has 10 keels; and (iv) pronounced differences in morphometric values (see Table I).
| | | |
| Carapace: | ||
| -length | | |
| -anterior width | | |
| -posterior width | | |
| Metasomal segment I: | ||
| -length | | |
| -width | | |
| Metasomal segment V: | ||
| -length | | |
| -width | | |
| -depth | | |
| Vesicle: | ||
| -width | | |
| -depth | | |
| Pedipalp: | ||
| -Femur length | | |
| -Femur width | | |
| -Tibia length | | |
| -Tibia width | | |
| -Chelae length | | |
| -Chelae width | | |
| -Chelae depth | | |
| Movable finger: | ||
| -length | | |
Notes on the habitat of Tityobuthus ivohibe n. sp.
The vegetation of the zone where T. ivohibe was captured at 900 m consists of largely undisturbed lowland humid forest. The site is a few hundred meters from the northwestern edge of the Réserve Spéciale (RS) dIvohibe and falls within a continuous forested corridor between this reserve and the Réserve Naturelle Intégrale (RNI) dAndringitra to the north. In the lowland forest of the RNI dAndringitra, at approximately the same altitude as the site at Ivohibe, the dominante large or emergent trees consist of Sloanea rhodantha var. rhodantha (Elaeocarpaceae) and Canarium madagascariense (Burseraceae) and the canopy and understory trees of members of the Lauraceae, Myrtaceae, Violaceae (Rinorea), Monimiaceae (Tambourissa, Ephippiandra, and Decarydendron) (Lewis et al. 1996).
In the vicinity of the 900 m site at the edge of the RS dIvohibe, down large trees, in various stages of decay, were relatively common and soil litter or the organic layer was not extensive. During the inventory of this site, little rain fell and the upper soil layer was distinctly dry.
The intent of the 1997 inventory of the forested Ivohibe region was to assess the elevational distribution of plants and animals along a transect of the slopes of Pic Ivohibe and the importance of the forested corridor between the RS dIvohibe and the RNI dAndringitra. Sites were surveyed at 900 m (just outside the reserve), at 1200 and 1575 m within the reserve, and at 900 and 1200 m in the corridor. Both 1200 m sites surveyed on Pic Ivohibe contained numerous montane elements, and was distinctly different from the 900 m sites.
Tityobuthus ivohibe was only found at the 900 m just outside the reserve and not at any other of the sites surveyed. Further, this species was not recorded during an intensive four month inventory of the RNI dAndringitra in 1993, although similar pitfall techniques were used. We presume that this species is relatively uncommon, or at least difficult to capture using pitfall techniques, rather than having a very limited distribution.
There is a continuous band of extant forest that runs from Pic dIvohibe north to the RNI dAndringitra, and then in a northeasterly direction through the Ikongo Forest to the area surrounding the Parc National de Ranomafana. Although Tityobuthus ivohibe is currently only known from 900 m at the edge of the RS dIvohibe, we strongly suspect that it occurs in the lowland forest across this zone. This habitat type is severely threatened in the region, largely for slash and burn agricultural practices. If indeed this species is limited to lowland forest, a considerable portion of its habitat has already been destroyed.
Acknowledgements
Permits to conduct these inventories were kindly granted by the Direction des Eaux et Forêts and Association National pour la Gestion des Aires Protégées. To officials of these organizations we are grateful. The inventory of the Ivohibe Forest was in the context of a WWF integrated conservation and development project in the Andringitra/Ivohibe region and we benefited enormously by the aid of colleagues based in Ambalavao. The field study at the Forêt dAnkazomivady was financed by a grant from the John D. and Catherine T. MacArthur Foundation.
References
Lewis, B.A., P.B. Phillipson, M. Andrianarisata, G. Rahajasoa, P.J. Rakotomalaza, M. Randriambololona & J.F. McDonagh, 1996. A study of the botanical structure, composition, and diversity of the eastern slopes of the Réserve Naturelle Intégrale dAndringitra, Madagascar, pp. 24-72. In: Goodman, S. M., ed., A flora and fauna inventory of the eastern slopes of the Réserve Naturelle Intégrale dAndringitra, Madagascar: with reference to elevational variation. Fieldiana: Zoology, new series, N° 85. [ Links ]
Lourenço, W.R., 1993. A review of the geographical distribution of the genus Ananteris Thorell (Scorpiones: Buthidae), with description of a new species. Rev. Biol. Trop., 41(3): 697-701. [ Links ]
Lourenço, W.R., 1994. Scorpions (Chelicerata) de Colombie. VI. Quatre nouvelles espèces de Buthidae des regions amazonienne, sud-pacifique et de la cordillère orientale. Rev. Acad. Colomb. Cienc., 19 (73): 387-392. [ Links ]
Lourenço, W.R., 1996. Scorpions. In: Faune de Madagascar. N 87. Muséum national dHistoire naturelle, Paris: 102 pp. [ Links ]
Lourenço, W.R., 1997. Another new species of Tityobuthus from Madagascar (Scorpiones, Buthidae). Entomol. Mitt. zool. Mus. Hamburg, 12 (155): 147-151. [ Links ]
Lourenço, W.R., 1998. Description of a new species of scorpion from the Réserve Spéciale dAnjanaharibe-Sud, Madagascar, pp. 69-72. In: Goodman, S.M., ed., A floral and faunal inventory of the Réserve Spéciale dAnjanaharibe-Sud, Madagascar: with reference to elevational variation. Fieldiana: Zoology, new series, N 90. [ Links ]
Lourenço, W.R. & S. M. Goodman (in press). Taxonomic and ecological observations on the scorpions collected in the Réserve Naturelle Intégrale dAndohahela, Madagascar. Fieldiana: Zoology, new series. 94: 149-153. [ Links ]
Vachon, M., 1963. De l'utilité, en systématique, d'une nomenclature des dents des chélicères chez les Scorpions. Bull. Mus. natn. Hist. nat., Paris 2è sér., 35(2): 161-166. [ Links ]
Vachon, M., 1973. Etude des caractères utilisés pour classer les familles et les genres de Scorpions (Arachnides). 1. La trichobothriotaxie en arachnologie. Sigles trichobothriaux et types de trichobothriotaxie chez les Scorpions. Bull. Mus. natn. Hist. nat., Paris, 3è sér., n 140, Zool., 104: 857-958. [ Links ]
Vachon, M., 1975. Sur l'utilisation de la trichobothriotaxie du bras des pédipalpes des Scorpions (Arachnides) dans le classement des genres de la famille des Buthidae Simon. C.R. séan. Acad. sci., Paris, sér. D, 281: 1597-1599. [ Links ]
Voss, R.S. & L.H. Emmons, 1996. Mammalian diversity in Neotropical lowland rainforests: a preliminary assessment. Bulletin of the American Museum of Natural History, 230: 1-115. [ Links ]

