










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.47 n.3 San José Sep. 1999
Abstract
This paper describes the fossils, materials and paleoenvironmental conditions found in some outcrops near the town of Jesús María, Turrialba, Costa Rica. The rock materials (reefal limestone, sandstones and conglomerates) are associated to the Punta Pelada Formation of Upper Oligocene to Lower Miocene age. Outcrops are of reduced extent and are unevenly distributed, hence they are considered as patch reefs developed by environmental energy fluctuations, sea level changes and clastic sedimentation from sources on small islands acting together in narrow platforms. The structural and environmental characteristics where fossil communities developed were derived from the analysis of 460 samples containing 36 species. The reefs represent low diversity communities comprising four coral species (three of them reef builders), 31 mollusks species (21 gastropods: a new species, 14 carnivores, three herbivores and three of uncertain feeding habits; 10 bivalves species: five endobenthic and five epibenthic); algae ( at least three groups represented); sea urchins; foraminifers; crabs. The evenness index is low, mainly regarding the Scleractinian corals, where Antiguastrea cellulosa predominates (80% of the recognized forms). The fossil assemblage suggests that the communities developed in shallow marine environments (50-80 m depth), affected by sea level changes, strong marine currents and clastic imput from nearby small islands.
Key words
Fossils, Systematics, corals, mollusks, Upper Oligocene, Lower Miocene, Costa Rica.
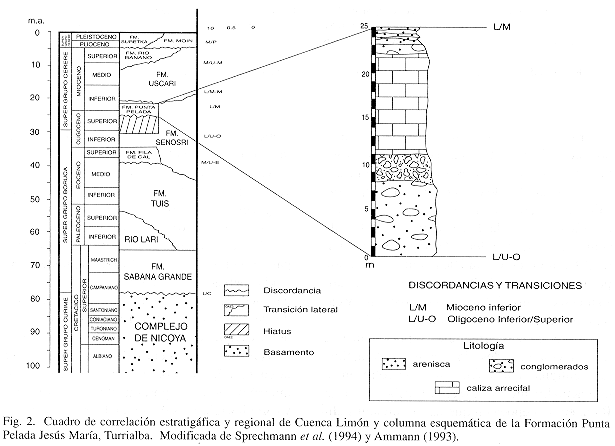
En la localidad de Jesús María, 4 km al noreste de la ciudad de Turrialba (Fig. 1), afloran de 12-30 m de calizas arrecifales, areniscas y conglomerados de edad Oligoceno Superior-Mioceno Inferior. Estas rocas se asocian estratigráficamente a la Formación Punta Pelada (Fig. 2), la cual fue definida originalmente para el Pacífico de Costa Rica como constituída por una secuencia de 225 m de areniscas, caliza arenosa, caliza lutítica, conglomerados y calizas arrecifales (Sprechmann et al. 1994).
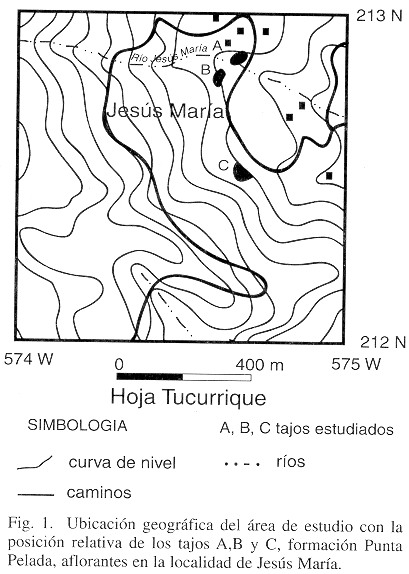
Estas calizas arrecifales representan una de las comunidades de corales Scleractinia más antiguas que inician su desarrollo en lo que actualmente es la región Caribe de Costa Rica, en una época en que las condiciones climáticas y geológicas no fueron las óptimas para el establecimiento y desarrollo de ese tipo de comunidades. Además, son un documento valioso para el estudio de los arrecifes de edad Oligoceno/Mioceno, los cuales son poco conocidos en el registro geológico mundial.
Las condiciones geológicas de Costa Rica para esa época, con plataformas angostas, formadas debido al reciente levantamiento de zonas aledañas (islas), no permitieron el desarrollo de grandes arrecifes. Esto combinado con una excesiva sedimentación clástica, producto de una fuerte actividad volcánica, cambios en la energía del medio, con eventos fuertes que interrumpieron temporalmente el crecimiento y cambios relativos del nivel del mar, causados por los levantamientos tectónicos que afectaban el área.
Por lo tanto sólo se establecieron arrecifes muy pequeños o de parche (2-5 m de espesor, 50 m de extensión), los cuales muestran una distribución irregular, condicionada tal vez por el sustrato preexistente.
La discusión detallada de los aspectos geológico-estratigráficos será tema de otro artículo.
Materiales y métodos
Para realizar los estudios de detalle, se recolectaron los ejemplares representativos (460 individuos) de las diferentes capas, durante un período de cinco años, realizando al menos tres visitas de recolección por año, tratando de obtener la mayor variedad posible y los mejores ejemplares, debido a la rápida desintegración de los materiales en los molinos de la cantera. Gran parte del material se recolectó de las acumulaciones realizadas durante las labores de extracción, siempre que se pudo relacionar con la capa de procedencia. Este procedimiento podría alterar los resultados en cuanto a la abundancia relativa entre los diferentes grupos, pero permitió conservar en una colección, ejemplares que pronto serían destruídos por el molino. Los fósiles recolectados fueron limpiados y procesados en el laboratorio y se estudiaron con lupa y microscopio binocular.
Además, se tomaron muestras de las rocas, siempre que se presentaron cambios litológicos notables, con el fin de realizar secciones delgadas y pulidas para el análisis de facies y el estudio de microfósiles.
La mayoría de los restos se encuentran alterados y se preservan de manera diversa. En los corales el material original está sustituído y presentan recristalización. Los moluscos: los gastrópodos en su mayoría se conservan como moldes y los bivalvos, muchos como moldes, pero los péctenes conservan su esqueleto original. Los otros grupos como cangrejos, briozoos, balanídeos y algas conservan su esqueleto.
Resultados
Las rocas estudiadas constituyen secuencias de calizas arrecifales, calizas masivas, calizas bioclásticas, brechas calcáreas, areniscas y conglomerados con afloramientos de espesores entre 12 y 30 m, con fuertes variaciones laterales y poca extensión horizontal.
El análisis de secciones delgadas permite determinar que las rocas son packstones (biolititas, biomicritas), wackestones (bioesparitas y biomicroesparitas) de corales, algas rojas calcáreas y foraminíferos con porcentajes menores de equinodermos, crustáceos, briozoos y moluscos. Además, hay grainstones, conformados principalmente por corales con restos de algas, moluscos y foraminíferos (James 1983). Son parasecuencias tipo CU (granocrecientes o coarsening upward), que se inician con una facies arrecifal (biolititas), la cual en algunas partes pasa a una facies lagunar o a facies de canal con areniscas y conglomerados rellenando canales de incisión, debido a la presencia de eventos de alta energía, asociados con tormentas. Sobre éstas se instalan de nuevo parches arrecifales, pasando en la parte superior de la secuencia a conglomerados con gradación normal, posiblemente asociados a fan deltas por retrabajo de los acantilados debido a un cambio del nivel del mar (lo cual se infiere de la organización interna de los conglomerados y de la presencia de fauna marina entre los clastos gruesos)..
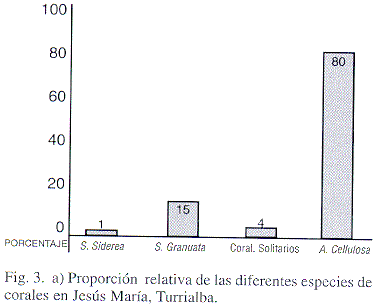
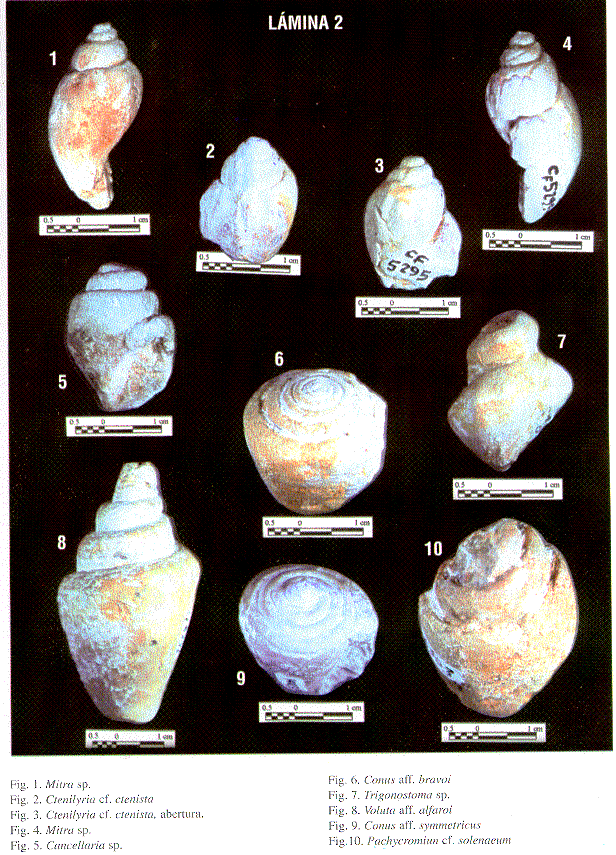
El estudio detallado de los macrofósiles, permitió determinar la presencia de 21 especies de gastrópodos (incluyendo el posible hallazgo de una especie nueva) de las cuales tres especies son herbívoras, 17 carnívoras y una de hábitos alimentarios desconocidos (Cuadro 1). Además, 10 especies de bivalvos, la mayoría de hábito epibentónico, aunque se presentan individuos endobentónicos asociados a los sedimentos y perforadores en los corales (Cuadro 1). Se reconocieron tres especies de corales hermatípicos, con un predominio marcado de Antiguastrea cellulosa (Fig. 3a). En algunas capas abunda una especie de coral solitario.
| Nombre | | |||||
| | | | | | | |
| Familia Turbinidae | ||||||
| Género Astraea RÖDING, 1798 | | |||||
| Astraea (Astralium) cf. brevispina basilis OLSSON | | |||||
| Familia Naticidae | ||||||
| Subfamilia Naticidae | ||||||
| Género Natica SCOPOLLI, 1777 | ||||||
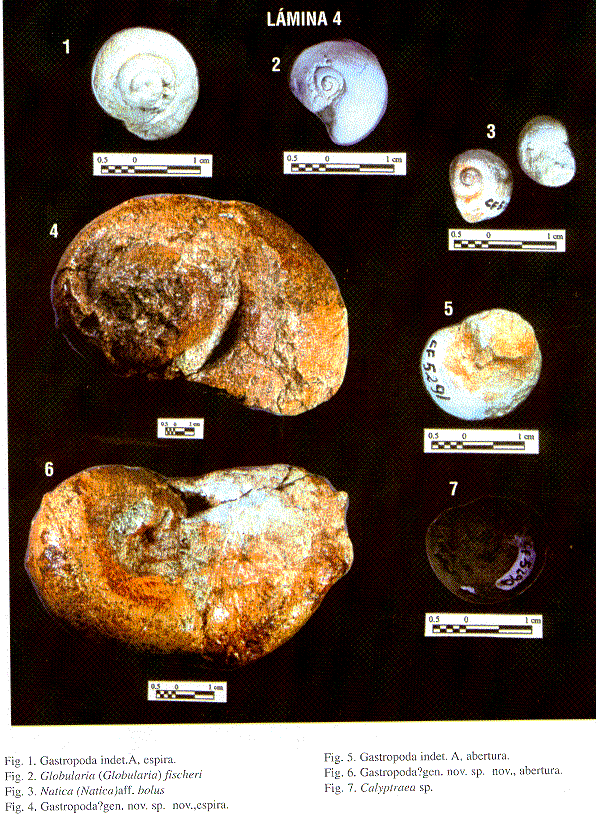
| Natica (Natica) aff.bolus Brown & PILSBRY | | |||||
| Subfamilia Globulariinae | ||||||
| Género Globularia SWAINSON | ||||||
| Globularia (Globularia) fischeri (DALL) | | | ||||
| Familia Ampullinae | ||||||
| Género Pachycromiun | ||||||
| Pachycromiun cf. solenaeum | | | ||||
| Familia Calyptraeidae | ||||||
| Género Calyptrea LAMARCK | ||||||
| Caliptraea sp. | | | | | ||
| Caliptraea (Trochita) trochiformis | | | | | ||
| Cypraea LINNAEUS | ||||||
| Cypaea sp. | | | | | ||
| Familia Melongidae | ||||||
| Melongena sp. | | | | |||
| Familia Fasciolariidae | ||||||
| Género Fasciolaria LAMARCK | ||||||
| Fasciolaria sp. | | | | |||
| Familia Volutidae | ||||||
| Subfamilia Lirinae | ||||||
| Género Ctenilyria WOODRING | ||||||
| Ctenilyria cf ctenista WOODRING | | | ||||
| Voluta aff alafaroi DALL | | | ||||
| Familia Xancidae | ||||||
| Género Xancus RÖDING | ||||||
| Xancus sp. | | | | | ||
| Familia Olividae | ||||||
| Género Oliva | ||||||
| Oliva sp. | | | | | | |
| Familia Mitridae | ||||||
| Género Mitra LAMARCK | ||||||
| Mitra sp. | | |||||
| Familia Conidae | ||||||
| Género Conus | ||||||
| Conus aff. bravoi SPIEKER | | |||||
| Conus aff. symmetricus SOWERBY | | | ||||
| Familia Cancellariidae | ||||||
| Género Cancellaria LAMARCK | ||||||
| Cancellaria sp. | | | | | ||
| Género Trigonostoma BLAINVILLE | | | | |||
| Trigonostoma sp. | ||||||
| Gastropoda indet. A | ||||||
| Gastropoda indet. B | ||||||
| Gastropoda ? gen. nov. sp. nov. | ||||||
| Familia Arcidae | ||||||
| Género Anadara GRAY | ||||||
| Anadara sp. | | | | | ||
| Familia Glycymeris | ||||||
| Género Glycymeris DA COSTA | ||||||
| Glycymeris sp. | | | | | ||
| Familia Mytilidae | ||||||
| Género Lithophaga RÖDING | ||||||
| Lithophaga sp. | | | | |||
| Familia Ostreidae | ||||||
| Género Ostrea Linnaeus | ||||||
| Ostrea sp. | | | | | | |
| Género Crassostrea SACCO | ||||||
| Crassostrea sp. | | | ||||
| Hytissa aff. Haitensis (SOWERBY) Familia Pectinidae | | | | |||
| Género Argopecten MONTEROSATO | ||||||
| Argopecten aff. levicostatus (TOULA) | | | ||||
| Familia Tellinidae | ||||||
| Género Strigilla TURTON | ||||||
| Strigilla aff. carnaria (LINNE) | | | ||||
| Bivalvia indet. A | ||||||
| Bivalvia indet. B | ||||||
| Clase Antozoa | ||||||
| Subclase Zoantharia | ||||||
| Orden Scleractinia | ||||||
| Familia Faviidae | ||||||
| Familia Siderastreidae | ||||||
| Familia Pocilloporidae | ||||||
| Antiguastrea Cellulosa (DUNCAN) | | | | |||
| Siderastrea siderae (ELLIS & SOLANDER) | | | | |||
| Stylophora ? granulata DUNCAN | | | ||||
| Corales solitarios indet. | ||||||
| Filo Artropoda | ||||||
| Subfilo Crustácea | ||||||
| Cirripedia indet. | | | | | | |
| Decapoda indet. | | | | | | |
| Filo Echinodermata | ||||||
| Echinoidea indet. | | | | | | |
| Filo Bryozoa | ||||||
| Clase Gymnolaemata | | | | | | |
| Framinífera plantónicos | | | | | | |
| Miliólidos | | | | | | |
| Lepidocyclina | | | | |||
| Nummulites | | |||||
| Algae | | | | | | |
| Oncoides | | | | | | |
| Lithophylum | | | | | | |
| Archaelithothamium | | | | | | |
En general se puede decir que la fauna y flora analizadas se desarrollaron durante el Oligoceno-Mioceno Inferior (Cuadro 2).
| Nombre | | ||||||
| | | | | | | | |
| Familia Turbinidae | |||||||
| Género Astraea RÖDING, 1798 | | ||||||
| Astraea (Astralium) cf. brevispina basilis OLSSON | | | | ||||
| Familia Naticidae | |||||||
| Subfamilia Naticidae | |||||||
| Género Natica SCOPOLLI, 1777 | |||||||
| Natica (Natica) aff.bolus Brown & PILSBRY | | | |||||
| Subfamilia Globulariinae | |||||||
| Género Globularia SWAINSON | |||||||
| Globularia (Globularia) fischeri (DALL) | | | | ||||
| Familia Ampullinae | |||||||
| Género Pachycromiuns | |||||||
| Pachycromiun cf. solenaeum | | | | ||||
| Familia Calyptraeidae | |||||||
| Género Calyptrea LAMARCK | |||||||
| Caliptraea sp. | | | | | | ||
| Caliptraea (Trochita) trochiformis | | | | | | ||
| Cypraea LINNAEUS | |||||||
| Cypaea sp. | | | | | | ||
| Familia Melongidae | |||||||
| Melongena sp. | | | | | | ||
| Familia Fasciolariidae | |||||||
| Género Fasciolaria LAMARCK | |||||||
| Fasciolaria sp. | | | | | |||
| Familia Volutidae | |||||||
| Subfamilia Lirinae | |||||||
| Género Ctenilyria WOODRING | |||||||
| Ctenilyria cf ctenista WOODRING | | | | | |||
| Voluta aff alafaroi DALL | | | | | |||
| Familia Xancidae | |||||||
| Género Xancus RÖDING | |||||||
| Xancus sp. | | | | | | ||
| Familia Olividae Género Oliva | |||||||
| Oliva sp. | | | | | | ||
| Familia Mitridae Género Mitra LAMARCK | |||||||
| Mitra sp. | | | |||||
| Familia Conidae | |||||||
| Gñenero Conus | |||||||
| Conus aff. bravoi SPIEKER | | ||||||
| Conus aff. symmetricus SOWERBY | | | |||||
| Familia Cancellariidae | |||||||
| Género Cancellaria LAMARCK | |||||||
| Cancellaria sp. | | | | | | ||
| Género Trigonostoma BLAINVILLE | | | | | |||
| Trigonostoma sp. | | | |||||
| Gastropoda indet. A | | ||||||
| Gastropoda indet. B | | ||||||
| Gastropoda ? gen. nov. sp. nov. | | | |||||
| Familia Arcidae | |||||||
| Género Anadara GRAY | |||||||
| Anadara sp. | | | | | | ||
| Familia Glycymeris | |||||||
| Género Glycymeris DA COSTA | |||||||
| Glycymeris sp. | | | | | | ||
| Familia Mytilidae | |||||||
| Género Lithophaga RÖDING | |||||||
| Lithophaga sp. | | | | | |||
| Familia Ostreidae | |||||||
| Género Ostrea Linnaeus | |||||||
| Ostrea sp. | | | | | | | |
| Género Crassostrea SACCO | |||||||
| Crassostrea sp. | | | | | |||
| Hytissa aff. Haitensis (SOWERBY) | | | | | | ||
| Familia Pectinidae | |||||||
| Género Argopecten MONTEROSATO | |||||||
| Argopecten aff. levicostatus (TOULA) | | | | | | ||
| Familia Tellinidae | |||||||
| Género Strigilla TURTON | |||||||
| Strigilla aff. carnaria (LINNE) | | | | ||||
| Bivalvia indet. A | | | |||||
| Bivalvia indet. B | | | |||||
| Clase Antozoa | |||||||
| Subclase Zoantharia | |||||||
| Orden Scleractinia | |||||||
| Familia Faviidae | |||||||
| Familia Siderastreidae | |||||||
| Familia Pocilloporidae | |||||||
| Antiguastrea Cellulosa (DUNCAN) | | | | ||||
| Siderastrea siderae (ELLIS & SOLANDER) | | | | | |||
| Stylophora ? granulata DUNCAN | | | | ||||
| Corales solitarios indet. | | | |||||
| Filo Artropoda Subfilo Crustácea | |||||||
| Cirripedia indet. | | | | | | | |
| Decapoda indet. | | | | | | | |
| Filo Echinodermata | |||||||
| Echinoidea indet. | | | | | | | |
| Filo Bryozoa | |||||||
| Clase Gymnolaemata | | | | | | | |
| Framinífera plantónicos | | | | | | | |
| Miliólidos | | | | | | | |
| Lepidocyclina | | | | ||||
| Nummulites | | | |||||
| Algae | | | | | | | |
| Oncoides | | | | | | | |
| Lithophylum | | | | | | | |
| Archaelithothamium | | | | | | | |
Los ejemplares estudiados se encuentran en la Colección de fósiles de la Escuela Centroamericana de Geología bajo los códigos CF-3613, 3614, 3817, 3818, 3820, 3822-3824, 3827, 3849, 3850, 3865, 3979-3982, 5278-5307, 5310-5321.
Se presenta una determinación sistemática de los moluscos y corales, por lo menos hasta nivel de género. Con los otros grupos presentes (briozoos, cangrejos, erizos, algas etc.), se hará una determinación muy general, a nivel de grupo taxonómico superior.
Formas predominantemente herbívoras, comen algas microscópicas que sacan de las rocas con la rádula, algunos también comen esponjas.
Referencias: KEEN 1971: 355; KELLUM 1926: 25; OLSSON,1922: 334; WOODRING 1928: 413; WEISBOARD 1962: 98; JUNG 1969: 423; MAURY,1917: 154.
Material: 6 ejemplares casi completos.
Descripción: Concha moderadamente grande, cónica con una base plana a algo cóncava. Espira alta, compuesta por cinco vueltas, las primeras no se preservan. Sutura ondulada y algo deprimida. Vueltas cóncavas y aplanadas hacia la sutura posterior.
Presentan indicios de pliegues diagonales que sobresalen en el contacto de las vueltas formando pústulos. Además, una periferia muy ondulada y con fuertes líneas espirales en la base.
Dimensiones:
Alto inc.: De 29 mm a 22mm
Ancho: De 49mm a 40 mm
Hábitat: Eulitoral medio-superior, zona rocosa.
Son en su mayoría de hábito hervíboro, sin embargo el género Natica es una excepción, es carnívoro, depreda activamente otros moluscos, principalmente bivalvos. Además, algunas especies de este órden son parásitos.
Referencias: KEEN 1971: 473; WOODRING 1957: 84; MAURY 1925: 238.
Material: 31 ejemplares, preservados principalmente la espira y parte de la habitación, falta la abertura.
Descripción: Semicircular u oval, con una espira baja y habitación grande. Superficie lisa. Hombros algo inflados.
Abertura grande, circular, pero no se puede determinar bien el ombligo debido a que está relleno de sedimento.
Dimensiones:
Alto: De 15mm a 7 mm
Ancho: De 18mm a 7mm
Hábitat: Ambiente marino somero (hasta 90m).
Referencias: WOODRING 1957: 94; 1959: 158; 1973: 475.
Material: Un ejemplar que presenta la abertura rellena de sedimento.
Descripción: Moderadamente grande, muy inflada, espira baja. No se puede observar la parte interna de la abertura ni la ornamentación. El molde interno es liso, con un surco bien marcado bordeando el labio interno.
Dimensiones:
Alto: 30mm
Ancho: 29mm
Discusión: Se parece mucho a la especie definida por Woodring 1959: 158, como Globularia aff. (Globularia) fischeri, pero no tiene la espira tan alta, por lo que se incluye en la especie G. fischeri. Con G. megista Woodring 1973: 475, difiere por ser de tamaño menor. Globularia es un género común en el Eoceno.
Referencias: WOODRING 1928: 391; 1957: 96; 1959: 158.
Material: Tres ejemplares con parte de la espira y de la habitación truncadas.
Descripción: Gastrópodos de tamaño medio, espira moderadamente alta, inflada, vueltas con hombro visible. Abertura semiovalada, corta.
Dimensiones:
Alto: 38mm incompleto
Ancho: 30mm
Discusión: La familia incluye muchos taxones fósiles, pero existe sólo una especie reciente.
El género Pachycromiun, ha sido reportado en capas del Eoceno, Oligoceno y Mioceno. Es muy común en el Eoceno, menos abundante en el Oligoceno y sobrevive hasta el Mioceno en el Golfo de México y la región Caribe.
Los ejemplares se relacionan con la especie P. solenaeun, descrita por Woodring 1957: 96, para el Eoceno Medio de la zona del Canal, Panamá.
Referencias: KEEN 1971: 455
Material: Un molde interno.
Descripción: Se preserva el ápice de conchas cónicas de tamaño medio, las cuales se pueden comparar al género Calyptraea.
Referencias: KEEN 1971: 456; WOODRING 1957: 81.
Material: Dos ejemplares completos y dos fragmentos.
Descripción: Concha cónica de tamaño medio, espira moderadamente alta. Apice truncado, central y espiral. La escultura consiste de fuertes pliegues axiales ablicúos. Abertura basal. El material se encuentra deformado.
Dimensiones:
Alto: 29mm
Ancho: 16 mm
Hábitat: Sobre rocas y detrito.
Discusión: Muy parecido a las formas descritas para el Oligoceno de Panamá, por Woodring 1957: 81.
Referencias: KEEN 1971: 492; INGRAM 1947: 5; WOODRING 1959: 194.
Material: Cuatro moldes internos con perforaciones.
Descripción: Concha pequeña, subredondeada. Abertura estrecha y rellena con una matriz muy fina, por lo que no se pueden observar las estrías. Labio externo curvo hacia el ápice. Superficie lisa.
Dimensiones:
Alto: 23mm
Ancho: 12mm
Hábitat: Común en aguas poco profundas.
Discusión: No se puede determinar la especie, debido a la mala preservación.
Es comparable con C. gurabensis Ingram, en la forma externa y el tamaño, pero no se pueden observar los dientes ni su disposición en la abertura.
Con pocas excepciones, los Neogastrópodos son carnívoros. Para lo cual presentan adaptaciones en el cuerpo blando y especialmente en la rádula.
Referencias: KEEN 1971: 604; WOODRING 1973: 478.
Material: Fragmento de la espira de un ejemplar grande.
Hábitat: Se encuentra al nivel de la marea baja.
Descripción: Espira de tamaño medio, con sutura muy constricta y con una especie de crenulaciones alrededor de las suturas.
Discusión: Se asemeja a M. propatula del Mioceno, pero la determinación es imprecisa por contarse solo con un ejemplar incompleto.
Hábitat: Viven generalmente cercanos a los manglares, en fondos fangosos, sobre los pisos mesolitoral, infralitoral y circalitoral y en la zona litoral en general.
Referencia: KEEN 1971: 611; OLSSON 1922: 278.
Material: Fragmentos de la habitación y partes de la espira, preservado como molde interno.
Descripción: Concha muy grande. La habitación tiene un hombro muy amplio, inclinado. Presenta prominencias a manera de várices o tubérculos en el borde del hombro. Por ser un molde interno y estar incompleto, no se puede observar ni la vuelta ni el canal sifonal.
Dimensiones:
Alto: 100mm
Ancho: 110mm ancho deformado
Hábitat: Las especies de este grupo viven sobre fondos arenosos, detríticos e icluso rocosos del piso infralitoral (Sbelli 1982).
Referencias: KEEN 1971: 619; WOODRING 1964: 288.
Material: Dos ejemplares, uno casi completo y uno fragmentado.
Descripción: Tamaño medio, bicónica, turriforme. La escultura axial consiste de pliegues fuertes, ampliamente espaciados, inclinados hacia adentro, hacia la sutura.
No se observa la ornamentación espiral y la abertura está rellena, por lo que no se pueden apreciar las denticulaciones del labio externo ni los pliegues de la columela.
Dimensiones:
Alto: A;27mm incompleto. B; 22.1mm
Ancho: A; 21mm B;15.2mm
Discusión: Los pliegues axiales fuertes, ampliamente espaciados y el contorno turriforme, tipifican el género. Aunque los ejemplares estudiados son de tamaño mayor. El género fue definido para el Eoceno de la Zona del Canal.
Referencia: OLSSON 1922: 271.
Material: Un ejemplar casi completo.
Descripción: Concha fuerte, alta, vuelta típicamente con hombro y doce pliegues, las vueltas son mas angostas y altas en el ángulo del hombro. Las vueltas parecen muy lisas. No se pueden observar estrías espirales debido a la preservación.
Dimensiones:
Alto: 56 mm
Ancho: 30 mm
Referencias: KEEN 1971: 620; WOODRING 1964: 285; 1973: 479.
Material: Fragmento de la habitación y molde interno de parte de la espira.
Descripción: A pesar de ser un fragmento, en las vueltas se nota una inclinación fuerte de la sutura al hombro. La habitación es subcilíndrica.
Discusión: Xancus es un género que vivió tanto en aguas del Atlántico Oeste, como del Pacífico Este, pero actualmente sobrevive sólo en el Caribe.
A pesar de ser un fragmento, el declive que presenta en el hombro y la forma de la habitación, permiten reconocer el género.
Se incluye el ejemplar en el género Xancus, a pesar de que la Comisión Internacional de Nomenclatura Zoológica (ICZN), en su opinión 489, 1957 (según Keen 1971: 620; Woodring 1964: 285), elimina este nombre en favor del nombre Turbinella. Sin embargo, el nombre Xancus tiene prioridad y ha sido ampliamente usado en fósiles del Eoceno, Oligoceno y Mioceno de la región.
Referencias: KEEN 1971: 622; PILSBRY & JOHNSON 1917: 165; BEAL,1948: 5; WOODRING 1964: 277.
Material: 28 ejemplares casi completos.
Descripción: Concha cilíndrica, de tamaño medio, algo ancha. Abertura larga, 3/4 del largo total de la concha, sutura acanalada angosta, con una espira baja formada por las primeras dos vueltas. Columela con pliegues, apenas visibles por el relleno de la abertura.
Dimensiones:
Alto: De 24mm a 15 mm
Ancho: De 15mm a 8mm inc.
Abertura: 2mm
Discusión: Los ejemplares aunque en su mayoría completos, no pueden compararse con las especies descritas en la literatura, debido a que son más anchas y de contorno más deprimido. Esto puede ser un efecto de la compactación, visible también en otros especímenes.
Hábitat: Viven en los pisos infralitorales o mesolitorales escondidos bajo el fango.
Referencias: KEEN 1971: 639
Material: Un ejemplar sin el ápice y con la abertura fragmentada.
Descripción: Concha de tamaño medio, lisa,espira alta, esbelta.
Hábitat: Se distribuyen generalmente en la zona fital, y muy raramente en la zona afital.
Referencias: KEEN 1971: 660; PILSBRY & JOHNSON 1917: 159; WOODRING 1970: 348.
Material: Seis fragmentos en su mayoría preservando la espira y uno con 3/4 partes de la habitación preservada.
Descripción: Concha típicamente cónica, ancha en la parte superior, en el hombro y angosta hacia la parte más anterior. Abertura larga, angosta y de ancho muy uniforme.
Buena preservación pero como molde, por lo que no se observa ningún tipo de ornamentación, que permita una determinación más precisa. Tamaño medio, hombros bien marcados, espira baja.
Dimensiones: Ejemplares incompletos.
Alto: De 36mm a 18mm
Ancho: De 19mm a 8mm
Hábitat: Generalmente aguas someras.
Discusión: Es similar a C. bravoi, pero de tamaño menor. Además, debido a la preservación, no se puede saber si presentan en el hombro las crenulaciones características de la especie.
Referencia: OLSSON 1922: 215; MUARY 1917: 206.
Material: Un ejemplar incompleto.
Descripción: Concha de tamaño medio, espira baja, mas o menos plana, hombros angostos. Presenta restos de estrías espirales prominentes en la habitación.
Dimensiones:
Alto: incompleto 20mm
Ancho: máximo 12mm
Referencias: KEEN 1971: 647
Descripción: Se asemeja en la forma externa, aunque el ejemplar estudiado es un molde.
Hábitat: Viven en los pisos infralitorales y circalitorales.
Referencias: KEEN 1971: 656; WOODRING 1928: 224
Material: Tres ejemplres fragmentados preservados como molde interno.
Descripción: Concha de tamaño medio, con sutura profunda, abertura ovalada. La escultura consiste de pliegues axiales fuertes y retractivos apenas visibles en el molde.
Dimensiones:
Alto: incompleto 20mm
Ancho: incompleto 10,5mm
Cuatro fragmentos de la espira y parte de la habitación de gastrópodos pequeños, preservados como moldes internos, muy deformados. Se puede apreciar una espira alta y delgada y una habitación grande y algo inflada.
Varios fragmentos de espirales de moluscos grandes, que no permiten reconocer el género.
Referencias: WOODRING 1959: 158; JUNG 1974: 496
Material: Un ejemplar preservado principalmente como molde interno.
Descripción: Gastrópodo de gran tamaño, con hombros bien marcados. Espira baja en comparación con el diámetro de la habitación. Vueltas de perfil convexo, sutura deprimida. Abertura extendida, de contorno subovalado. Ombligo amplio. Parte basal amplia y plana.
Dimensiones:
Alto: 72 mm
Ancho: 160 mm
Discusión: No se encontraron formas parecidas en la literatura revisada, lo más aproximado es el género Globularia (Woodring 1957, 1959), sin embargo el ejemplar estudiado es de gran tamaño, tiene una base muy amplia y el ombligo muy ancho. Jung (1974) describe un ejemplar como Naticidae indet., que es comparable en tamaño con el ejemplar estudiado, pero difiere de éste por presentar la habitaciòn y la abertura más alargadas. Se considera que puede representar un género y especie nuevos, pero por no contar con una muestra más representativa, no se hace la descripción formal.
Charnela con muchos dientes, contorno trapezoidal, escultura radial.
Material: Molde interno de la valva izquierda.
Descripción: Concha alargada, umbón subcentral ubicado en el cuarto anterior.
Dimensiones:
Alto: 10mm
Ancho: 23mm
Discusión: La forma externa y la ubicación del umbón son los únicos criterios que permiten incluir el ejemplar en el género Arca.
Hábitat: En fondos arenosos o lodosos a profundidad variable.
Referencias: KEEN 1971: 54
Material: Alrededor de treinta ejemplares, la mayoría preservados como moldes internos, muy pocos con restos de la concha con escultura. Pero siempre se encuentran las valvas conjugadas.
Descripción: Concha subcircular, equivalva, umbón central. Escultura constituida por estrias radiales finas.
Dimensiones:
Alto: De 9mm a 20mm
Ancho: De 10mm a 25mm
Hábitat: Aguas someras.
Conchas equivalvas, fuertemente inequilaterales, las impresiones de los músculos aductores de diferente tamaño.
Referencias: Keen 1971: 68
Material: Tres ejemplares completos, y varios tubos de la perforación con el bivalvo adentro.
Descripción: Concha de tamaño medio, cilíndricas con estrías de crecimiento bien maracadas en el lado posterior.
Dimensiones:
Alto: De 18mm a 34mm
Ancho: De 9mm a 11 mm
Hábitat: Perforando rocas o conchas.
Generalmente inequivalvas y algo inequilaterales; concha de material prismático-nacarado a foliáceo; línea integripaleada.
Referencias: Keen 1971: 82
Material: Varios fragmentos de concha muy gruesa, y grande.
Descripción: Conchas compuesta por láminas. En algunas se puede observar el area ligamental ancha en forma más o menos triangular, con una depresión central triangular (resilífero) para el ligamento; en algunos ejemplares se puede apreciar una impresión muscular grande y circular.
Hábitat: Sobre sustratos duros, en aguas someras.
Referencias: WOODRING 1982: 610.
Material: 3 valvas de ejemplares pequeños.
Descripción: Valvas de tamaño pequeño, ovaladas verticalmente, moderadamente gruesas, espatuladas, laminación concéntrica uniforme o casi ausente. Debido a la preservación, no se puede observar el interior de las valvas.
Dimensiones:
Alto: 21 mm
Ancho: 11 mm máximo
Discusión: Los especímenes presentan la forma típica del género, aunque son de tamaño muy pequeño comparado con las descripciones presentadas para especies del género (p.e. C. cahobasensis (Pilsbry & Brown) in Woodring 1982: 610.
Hábitat: Aguas someras.
Referencias: KEEN 1971: 84; WOODRING 1982: 607; FISCHER 1985: 199.
Material: Una valva derecha bien preservada.
Descripción: Valva de tamaño pequeño, gruesa, subcircular, algo convexa, con cinco pliegues radiales fuertes, altos, redondeados, más visibles hacia la parte anterior marginal. Presenta láminas concéntricas, las cuales al intersectar los pliegues formas como pústulos.
Dimensiones:
Alto: 20 mm
Ancho: 25 mm
Discusión: El ejemplar estudiado parece ser un especímen juvenil de Hyotissa haitensis, por lo que no presenta las típicas espinas huecas de la especie, sólo hay prominencias a manera de nódulos.
Hábitat: El género Hyotissa actualmente habita arrecifes coralinos.
Referencias: OLSSON 1922: 371; FISCHER & FRANCO 1979: 37; WOODRING 1982: 596; KEEN 1971: 87.
Material: Alrededor de 20 ejemplares, la mayoría representados por valvas sueltas, dos de ellos con las valvas conjugadas.
Descripción: Conchas pequeñas, alto y ancho aproximadamente iguales, convexidad algo diferente en las dos valvas, aurículas casi iguales. Ornamentación consistente de pliegues radiales (19-25) altos, de cresta más o menos agudas,separadas por interespacios redondeados y profundos. Presentan láminas concéntricas finas, las cuales son más visibles en los ejemplares más pequeños. Las aurículas presentan escultura radial (6 pliegues), la cual es un poco más fuerte en la anterior derecha. La incisión bisal es poco profunda.
Dimensiones:
Alto: 11 mm máximo
Ancho: 12 mm máximo
Discusión: Los ejemplares estudiados coinciden en todo excepto en el tamaño con las formas descritas por Woodring 1982: 596; de la descripción de Fischer & Franco 1979: 37, difieren por tener un tamaño menor, por presentar laminaciones concéntricas y ambas aurículas ornamentadas. Debido a su semejanza, se considera una especie antecesora de la especie A. laevicostatus.
Hábitat: Aguas someras.
Conchas equivalvas, charnela heterodonta, con dos impresiones musculares.
Referencias: WOODRING 1982: 666; KEEN 1971: 233.
Material: Treinta moldes internos de las valvas conjugadas.
Descripción: Conchas pequeñas, triangulares-ovaladas, inequilaterales, el lado posterior algo más extendido que el anterior. Convexas, el lado anterior redondeado. Uno de los ejemplares presenta restos de una escultura fina divergente.
Dimensiones:
Alto: De 8 mm a 18 mm
Ancho: De 4.9 mm a 8.5 mm
Diámetro: De 18 mm a 9 mm
Hábitat: Enterradas en fondos blandos.
Discusión: Parece ser una especie precedente a Strigilla carnaria.
Dos moldes internos de bivalvos de tamaño medio, en los cuales no se puede observar ningún rasgo que permita hacer una identificación precisa.
Alrededor de ventiún especímenes de moldes internos de bivalvos, muchos de ellos con valvas conjugadas, de tamaño pequeño, convexas. Debido a la preservación no se pueden determinar ni a nivel genérico.
Referencia: VAUGHAN-WAYLAND 1919; WEISBORD 1973.
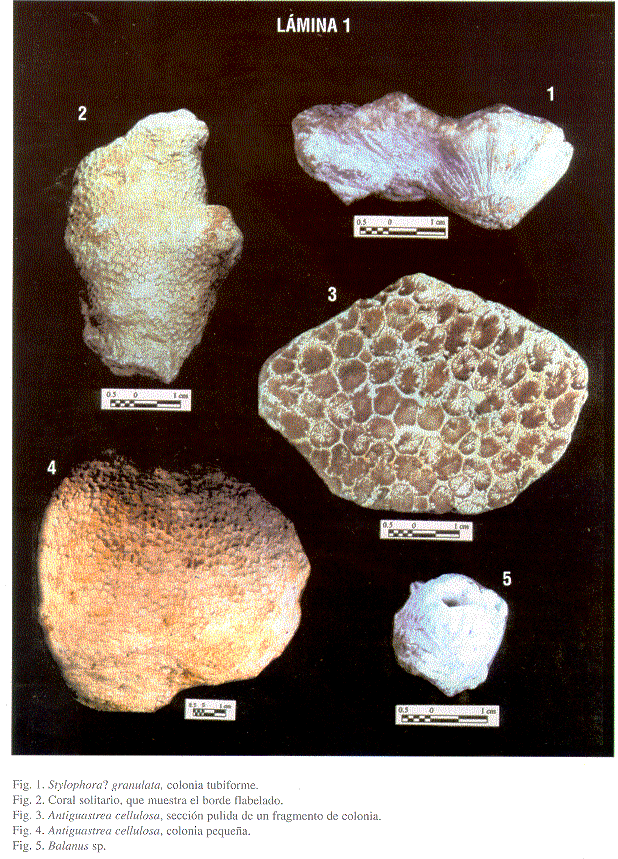
Material: Es la especie más abundante, con colonias de coral que varían de 5 hasta 70 cm de diámetro.
Descripción: Coralitos muy numerosos, altos, delgados, apiñados pero diferenciables, generalmente cilíndricos, aunque algunas veces son más o menos prismáticos. Tamaño variable, generalmente los cálices tienen un diámetro entre 2,5 y 4 mm. La sección transversal de los coralitos es subcircular, son algo profundos. Los septos se presentan en 4 ciclos, los primarios son los más largos, los secundarios son casi del mismo largo que los primarios, los terciarios y cuaternarios son más pequeños y más díficiles de observar. Los septos son por lo general retos, la columela es pequeña y poco desarrollada. La pared parece ser fuerte y tanto la endoteca como la exoteca están bien desarrolladas.
Observaciones: En algunos ejemplares se pueden observar los coralitos separados por áreas de alrededor de 1 mm de ancho.
Distribución Estratigráfica: Oligoceno- Mioceno Medio.
Referencias: VAUGHAN-WAYLAND 1919; WEISBORD 1973.
Material: Muy pocos ejemplares de tamaño medio, muy recristalizados.
Descripción: Los coralitos son altos, moderadamente delgados. Se pueden observar cálices irregularmente poligonales, con diámetros entre 2 mm y 3 mm, separados por paredes angostas, rectas. Los septos son gruesos, se pueden observar 4 ciclos, los primarios y secundarios se prolongan hasta la columela (aunque los primarios son un poquito más largos), los terciarios y cuaternarios son más cortos y no siempre se pueden diferenciar.
Columela pequeña, con una papila central.
Distribución Estratigráfica: Mioceno Inferior-Reciente.
Referencias: VAUGHAN-WAYLAND 1919.
Material: Numerosos coralarios ramosos, más o menos cilíndricos, con frecuencia presentan un lado aplastado. Los cálices se distribuyen irregularmente y están separados por un coenénquima granuloso. Son cálices circulares, profundos y rodeados por un anillo prominente de septos y costillas.
No se pueden diferenciar los septos ni la columela, debido a la preservación.
Distribución Estratigráfica: El género Stylophora es comunicado del Oligoceno, la especie S. granulata se reporta a partir del Mioceno Inferior.
En algunas de las facies se presentan muchos ejemplares de corales solitarios no atados a ningún sustrato, algo comprimidos, de tamaño pequeño (1-2.5 cm de alto). De forma cónica, ensanchando el diámetro fuertemente hacia el cáliz. El cáliz presenta un contorno superior flabelado, con un diámetro máximo de 5cm. Los septos sólo se pueden observar en los lados del coralito, son gruesos y regularmente espaciados. El cáliz se nota algo deprimido, pero debido a la preservación no se puede observar ni los tabiques, ni ninguna otra estructura del interior.
Se encuentran gran cantidad de restos de Balanídeos, como placas sueltas, como individuos aislados o en pequeñas agrupaciones. Los individuos son pequeños (1.3 cm de altura).
Se conservaron varios restos de tenazas de cangrejos, además de fragmentos del caparazón.
Se encuentran algunas colonias de briozoos formando costras sobre colonias de coral y ostras.
Se reconocen restos de esqueletos, generalmente muy fracturados y comprimidos y espinas de erizos irregulares.
En la secuencia estudiada se presentan áreas con grandes acumulaciones de foraminíferos, principalmente Lepidociclina, Miliólidos, algunos fragmentos de Nummulites, y otros restos de foraminíferos plantónicos no determinados.
Un constituyente muy importante de todas las secciones estudiadas son las algas. Son principalmente algas rojas coralinas del género Lithophyllum. En algunas partes se encuentran restos que parecen Archaeolithothamnium.Además, hay niveles con concentraciones de oncoides, los cuales en algunos casos recubren esqueletos de macroforaminíferos o fragmentos de otros organismos.


Paleoecología
Por encontrarse organismos en su ambiente natural (biocenosis) y restos transportados de ambientes vecinos (tafocenosis), se hicieron algunas deducciones sobre los factores ambientales que influyeron en la instalación, desarrollo y desaparición de las comunidades.
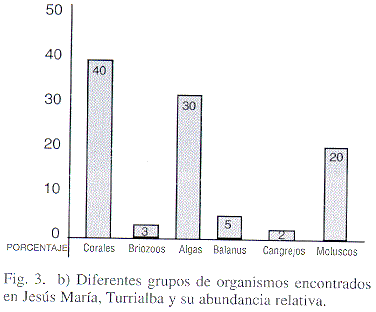
Son comunidades de baja diversidad (tres especies de constructores), en comparación con otros arrecifes del Oligoceno (14 géneros en Puerto Rico), o con arrecifes modernos del Caribe de Costa Rica (80 especies conocidas) (Fig. 3a,b). Por lo que se deduce que estas comunidades aprovecharon los lapsos de tiempo favorables para crecer.
La equidad también es baja, principalmente en relación con los corales, que presentan un marcado predominio de Antiguastrea celullosa (80% de las formas encontradas) (Fig. 3 a).
La baja diversidad y la baja equidad son indicadores de un desarrollo en un ambiente de poca profundidad, con influencia ocasional de corrientes fuertes y aporte de material terrígeno procedente de un área levantada en las cercanías.
Mediante el estudio de los organismos encontrados (460 individuos procesados y analizados), se determinó la presencia de al menos 36 especies : cuatro especies de corales (tres de constructores); 31 especies de moluscos (21 de gastrópodos: incluyendo el hallazgo de una especie nueva; 14 de carnívoros, tres de herbívoros, tres indeterminados; 10 especies de bivalvos: cinco endobentónicos y cinco epibentónicos), algas (representantes de por lo menos tres grupos), equinodermos, foraminíferos, crustáceos (Cuadro 1, 2).
Entre los macrofósiles destacan por su abundancia los corales (40%), las algas calcáreas (30%) y los moluscos (20%), en menor cantidad se encuentran restos de cirripedios, decápodos, echinoideos y briozoos (Fig. 3b).
El análisis de las microfacies indica un ambiente de depositación tipo lagoon con circulación abierta (shelf lagoon open circulation, según Flügel 1982), para las calizas (wackestones/ bioesparita y packestone/ biomicrita); áreas poco profundas con energía moderadamente alta para las calizas con oncoides, areniscas y conglomerados (Flügel 1982).
La fauna y la flora encontradas permitió deducir los factores ecológicos que influyeron para que se formara la estructura:
Luz: debido a la presencia de corales hermatípicos, los cuales requieren para su desarrollo la presencia de algas simbióticas asociadas y al hallazgo de muchos pectínidos, los cuales en muchas ocasiones tienen organelos fotosensibles, así como otros moluscos típicos del ambiente arrecifal o de fondos arenosos someros (Astraea, Oliva, Glycymeris), se interpreta que la asociación se desarrolló en la zona fótica.
Además, en toda la secuencia son importantes los restos de algas calcáreas, las cuales en algunas partes se encuentran formando oncoides (estructura calcárea, laminada concentricamente, formadas por la acreción de capas de algas), lo que confirma la intepretación de que las rocas se depositaron en la zona fótica a una profundidad variable, con aguas agitadas (Raup & Stanley 1978, Prager & Ginsburg 1989).
Salinidad: el contenido fosilífero (corales hermatípicos, equinodermos, briozoos, moluscos) permite inferir una salinidad normal (marina), lo cual es requisito indispensable para la instalación y el desarrollo de los arrecifes coralinos y muchos de los organismos asociados a ellos (echinodermos, briozoos) (Raup & Stanley 1978, Goreau et al. 1979).
Profundidad: las comunidades se desarrollaron en aguas marinas poco profundas (zona fótica), debido a la gran cantidad de algas calcáreas, las cuales muchas veces forman oncoides (profundidades menores de 80m, zona de alta turbulencia) y al crecimiento de colonias de corales (Raup & Stanley 1978).
Temperatura: se desarrollaron en aguas tropicales con temperaturas promedio superior a los 20°, (con un rango máximo permitido entre los 18°-23°C) en las que crecieron corales hermatípicos y otros organismos asociados, como algas, con esqueletos carbonatados (Vegas 1971, Andrews & Pickard 1990).
Turbulencia: las comunidades representadas se desarrollaron en el área de influencia de la turbulencia (hasta los 50-80 m). Esto se puede deducir de la gran cantidad de oncoides que se encuentran en algunos niveles (menos de 80m de profundidad). Además, en algunas partes se encuentran corales removidos, aunque no se evidencian marcas de desgaste por transporte en los restos conservados (Vegas 1971, Prager & Ginsburg 1989).
Sedimentación y turbidez: los parches se instalaron en una zona con influencia de sedimentación clástica, por lo que sólo se desarrollaron comunidades de tamaño reducido, con baja diversidad, rodeadas por sedimentos de granos fino. Además, es notable la destrucción en diversas ocaciones de estas comunidades, por el establecimiento de sedimentación clástica.
En las secuencias analizadas se pueden reconocer algunas épocas con sedimentación baja, evidenciadas por la instalación de corales del género Antiguastrea, llegando a veces a presentar altas tasas de bioerosión por Lithophaga, seguidas de épocas con un aumento en la sedimentación de grano fino, con predominio de especies con mayor capacidad de soportar altas tasas de sedimentación (formas ramosas del género Stylophora, bivalvos, algas), culminando con eventos de sedimentación siliciclástica.
Las secuencias pueden representar ciclos de progradación (ciclos CU), constituyendo en general una secuencia regresiva o puede evidenciar cambios en la energía del medio con la presencia de eventos de alta energía cíclicos o esporádicos.
Eh/Ph: las rocas carbonatadas se desarrollan en ambientes oxigenados lo que indica un Eh aproximado a 0.78 y un Ph entre 9-10 (Krauskopf 1967).
Sustrato: del análisis de las secuencias se puede deducir que el sustrato primario fue arenoso, estable y firme (en algunas partes blando), el cual fue colonizado por algas, cangrejos, gastrópodos y bivalvos endobentónicos. El sustrato secundario estuvo formado por la acumulación de fragmentos grandes de esqueletos, fue un sustrato duro, que permitió la colonización por organismos epilíticos y el reclutamiento de constructores. El sustrato firme fue colonizado por organismos endobentónicos, epibentónicos vágiles y hemisésiles (bivalvos con bisus y libres). Además, las algas colonizaron un sustrato duro secundario que formó el núcleo de los oncoides. La presencia de endolitismo (organismos que viven dentro del coral perforándolo, podría indicar que en partes los corales habían perdido el tejido orgánico, debido a lesiones en la superficie de las colonias (causado por organismos, por enfermedades o por fracturas), condición necesaria para que Lithophaga empiece a perforar, luego continúa dentro del esqueleto de la colonia viva (Fischer 1997 com. escrita).
En general se puede decir que las secuencias se depositaron en un ambiente de poca profundidad, desarrollándose pequeñas plataformas carbonatadas en una zona con influencia ocasional de corrientes fuertes y aporte de material terrígeno. El análisis de las microfacies indica un ambiente de depositación tipo Lagoon con circulación abierta (7: laguna de la plataforma de circulación abierta, según Flügel 1982) para las calizas (wackestones/bioesparita y packestones/ biomicritas) y áreas poco profundas con energía moderadamente alta para las calizas con oncoides, areniscas y conglomerados (Flügel 1982).
Se consideran arrecifes de parche debido a su tamaño pequeño (2-5 m de espesor) y a su distribución irregular, compuestos por comunidades de baja diversidad, que aprovecharon cortos lapsos para crecer. Estos arrecifes no se desarrollaron completamente, lo cual permite inferir la existencia de uno o varios de los siguientes factores:
-plataformas angostas, que no permitieron el desarrollo de grandes arrecifes
-excesiva sedimentación clástica producto de una fuerte actividad volcánica
-cambios en la energía del medio, con eventos que interrumpieron el crecimiento
-cambios del nivel del mar, tal vez debidos a levantamiento tectónico
Las rocas estudiadas representan biocenosis (arrecifes coralinos asociados a algas, moluscos, equinoideos y cangrejos) y tafocenosis (acumulaciones de restos transportados de un costa rocosa no muy lejana, como balánideos y ostras) de organismos que vivieron en ambientes marinos, tropicales y someros, las cuales son de baja diversidad y baja equidad.
Agradecimientos
El trabajo se realizó en el marco del proyecto de investigación N1 113-95-551, inscrito en la Vicerrectoría de Investigación de la Universidad de Costa Rica. La determinación taxonómica de los corales fue realizada por Nancy Budd (com. escrita 1993). Mis reconocimientos a J. Cortés, R. Fischer, P. Denyer, G. Alvarado, L.G. Salazar, A. Mata, S. Rojas, S. Hernández, W. Pérez y A. Loaiza por su valiosa colaboración en las diferentes etapas de realización de éste trabajo. A los revisores de este trabajo, por su paciencia y dedicación.
Resumen
En la localidad de Jesús María, Turrialba, afloran de 12-30 m de calizas arrecifales, areniscas y conglomerados de edad Oligocena Superior-Miocena Inferior, que se asocian con la Formación Punta Pelada.
En ésta localidad se registra una de las primeras comunidades arrecifales de la actual región Caribe de Costa Rica y constituye una de las pocas localidades de arrecifes de ésta edad en el área. Los afloramientos se interpretan como parches arrecifales debido a su distribución irregular y a su poca extensión lateral (50m), los cuales se desarrollaron bajo la influencia de diversos procesos: variación en la energía del medio, cambios en el nivel del mar, plataformas angostas, excesiva sedimentación clástica posiblemente desde islas.
Se analizaron 460 ejemplares, los cuales corresponden a más de 36 especies, que permitieron hacer una recosnstrucción de la estructura y de las condiciones ambientales en que se desarrollaron. Los arrecifes constituyeron comunidades de baja diversidad: cuatro especies de corales (tres de constructores), 31 especies de moluscos (21 de Gastrópodos: una especie nueva, 14 de carnívoros, tres de herbívoros, tres de hábitos alimenticios desconocidos; 10 especies de Bivalvos: cinco endobentónicos, cinco epibentónicos), algas (representantes de por lo menos tres grupos), equinodermos, foraminíferos, crustáceos. La equidad también es muy baja, principalmente en lo referente a los corales Scleractinia, con un predominio muy marcado de Antiguastrea celullosa (80% de las formas encontradas). Esto permite inferir que se desarrollaron en un ambiente de poca profundidad (50-80 m), muy variable, con influencia ocasional de corrientes marinas fuertes y aporte de sedimentos terrígenos procedentes de islas cercanas.
Referencias
Andrews, J. & G. Pickard. 1990. The Physical Oceanography of Coral-Reefs Systems. Coral Reefs Ecosystems of the World 25: 11-48. [ Links ]
Beal, C. 1948. Reconnaissance of the Geology and oil possibilities of Baja California, Mexico. Geol. Soc. Amer. Mem. 31: 1-138. [ Links ]
Fischer, R. 1985. La Fauna de la Formación Turrúcares (Mioceno, Valle Central, Costa Rica). Geol. Paleont. 19: 191-225. [ Links ]
Fischer, R. & J. Franco. 1979. La Formación Coris (Mioceno, Valle Central, Costa Rica). Inst. Geogr. Nal. Inf. sem. I-79:15-72. [ Links ]
Flügel, E. 1982. Microfacies analysis of limestones. Springer-Verlag, Berlín. 633. [ Links ]
Goreau, T., N. Goreau & T. Goreau. 1979. Corales y arrecifes coralinos. Invest. Cien. 37:48-60. [ Links ]
Ingram, W. 1947. Check list of the Cypraeidae occurring in the western Hemisphere. Paleontological Research Institution, Ithaca, Nueva York: 1-25. [ Links ]
James, N. 1983. Reef environment. Memoir-AAPG (33): 508-537. [ Links ]
Jung, P. 1969. Miocene and Pliocene Mollusks from Trinidad. Bull. Amer. Paleont. 247:293- 657. [ Links ]
Jung, P, 1974. Eocene Mollusks from CuraÇao, West Indies.En Jung P.: Contributions to the Geology and Paleontology of the Caribbean and Adjacent areas. Verhandl. Naturf. Ges. Basel. 84 (1):483-500. 7 lam. [ Links ]
Keen, M. 1971. Sea shells of tropical West America. Marine Mollusks from Baja California to Peru. Stanford University, Stanford, California 1964p. [ Links ]
Kellum, L. 1926. Paleontology and Stratigraphy of the Castle Hayne and trent marls in North Carolina. U. S. Geol. Surv. Prof. Paper 143:1-56. [ Links ]
Krauskopf, K. 1967. Introduction to Geochemistry. Mcgraw-Hill. Nueva York.721p. [ Links ]
Maury, C.J. 1917. Santo Domingo type sections and fossils. Bull. Am. Paleont., 5: 1- 477. [ Links ]
Maury, C.J. 1925. A further contribution to the Paleontology of Trinidad (Miocene horizons). Bull. Amer. Paleont. 42: 1-370. [ Links ]
Olsson, A.A. 1922. The Miocene of Northern Costa Rica. With notes on its general stratigraphic relations. Bull. Amer. Paleont. 9/39:173-481. [ Links ]
Prager, E. & R. Ginsburg. 1989. Carbonate nodule growth on Florida`s outer shelf and its implications for fosssil interpretations. Palaios 4:310-317. [ Links ]
Pilsbry, H. A. & C. W Johnson. 1917. New Mollusca of the Santo Domingan Oligocene. Procc. of the Acad. of Nat. Sci. of Philadelphia. 29:151-201. [ Links ]
Raup, D. & S. Stanley. 1978. Principios de Paleontología. Ariel, España. 456p. [ Links ]
Sbelli, B. 1982. Guía de Moluscos. Grijalbo, Barcelona.512p. [ Links ]
Sprechmann, P., A. Astorga, A. Fernández & C. Calvo. 1994. Cuadro sinóptico de correlación estratigráfica de las Cuencas Sedimentarias de Costa Rica. Rev. Profil 7: 427-431. [ Links ]
Vaughan-Wayland, T. 1919. Fossil Corals from Central America, Cuba, and Porto Rico, with an account of the American Tertiary, Pleistocene, and recent coral reefs. Bull. 103: 1-524. [ Links ]
Vegas, M. 1971. Introducción a la Ecología del Bentos Marino. Organización de Estados Americanos, Washington, D.C.98p. [ Links ]
Weisbord, N. 1962. Late Gastropods from northern Venezuela. Paleontological Research Institution Ithaca, Nueva York 42: 1- 672. [ Links ]
Weisbord, N. 1973. New and litle known corals from the Tampa Formation of Florida. Florida Dep. of Nat. Res. Bull.56: 1-146. [ Links ]
Woodring, W.P. 1928. Miocene Mollusks from Bowden, Jamaica. Carnegie Institute of Washington, DC. Washington. 564p. [ Links ]
Woodring, W.P. 1957. Geology and Paleontology of Canal Zone and Adjoinig Parts of Panama. Description of Tertiary Mollusks (Gastropods: Trochidae to Turritellidae). Geol. Surv. Prof. Paper 306-A:1-145, Lam. 3-23. [ Links ]
Woodring, W.P. 1959. Geology and Paleontology of Canal Zone and Adjoinig Parts of Panama. Description of Tertiary Mollusks (Gastropods: Vermetidae to Thaididae). Geol. Surv. Prof. Paper 306-B:147-239, Lam. 24-38. [ Links ]
Woodring, W.P. 1964. Geology and Paleontology of Canal Zone and Adjoinig Parts of Panama. Description of Tertiary Mollusks (Gastropods: Columbellidae to Volutidae). Geol. Surv. Prof. Paper 306-C:241-297, Lam. 39-47. [ Links ]
Woodring, W.P. 1970. Geology and Paleontology of Canal Zone and Adjoinig Parts of Panama. Description of Tertiary Mollusks (Gastropods: Eulimidae, Marginellidae to Helminthoglyptidae). Geol. Surv. Prof. Paper 306-D:299-452, Lam. 48-66. [ Links ]
Woodring, W.P. 1973. Geology and paleontology of Canal Zone and adjoining parts of Panama (Additions to gastropods, scaphopods, pelecypods: Nuculidae to Mallaeidae). Geol.Surv., Prof.Pap. 306-E:453-539, lám.67-82. [ Links ]
Woodring, W.P. 1982. Geology and paleontology of Canal Zone and adjoining parts of Central America. Description of Tertiary mollusks (Pelecypods:Propeamussidae to Cuspidariidae; additions to families covered in P-306-E; additions to gastropods; cephalopods). Geol.Surv., Prof.Pap. 306-F:541-759, lám.83-124. [ Links ]
1 Sección de Paleontología y Estratigrafía, Escuela de Geología, Universidad de Costa Rica, 2060 San José, Costa Rica. Fax (506) 234-2347.E-mail: aaguilar@cariari.ucr.ac.cr

