










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.47 n.3 San José Sep. 1999
Abstract
We compared the reproductive systems in populations of Clidemia capitellata (Bonpl.) D. Don, C. bullosa DC. and C. hirta (L.) D. Don. (Melastomataceae). The three species occur in small populations in forest margins at Mata de Dois Irm![]() os, Recife, Pernambuco, Brasil and are sympatric. They flower throughout the year, and are occasionaly visited by Halictidae bees, Augochloropsis sp.; the bees collect pollen by vibration ("buzz pollination"). The three species are agamospermous and exhibit a high level of male sterility, as measured by their low pollen viability.
os, Recife, Pernambuco, Brasil and are sympatric. They flower throughout the year, and are occasionaly visited by Halictidae bees, Augochloropsis sp.; the bees collect pollen by vibration ("buzz pollination"). The three species are agamospermous and exhibit a high level of male sterility, as measured by their low pollen viability.
Key words
"Buzz pollination", agamospermy, hibridization, Clidemia, Melastomataceae.
Los trabajos referentes al sistema reproductivo de Melastomataceae han demostrado la existencia de varias especies agamospérmicas que habitan diferentes ecosistemas de Suramérica (Baumgratz e Silva 1986, Renner 1989, Borges 1991, Goldenberg 1994, Melo & Machado 1996). Sin embargo, debido al escaso número de especies estudiadas, existen dudas razonables sobre el origen de la agamospermia en la familia.
El objetivo de nuestro trabajo fue investigar aspectos comparativos de fenología, biología floral, del sistema reproductivo y del comportamiento meyótico de las células madres del polen entre tres especies de Clidemia.
Materiales y métodos
Estudiamos las especies Clidemia capitellata (Bonpl.) D. Don., C. bullosa DC. (arbustos de 1,5 a 2,0m de altura) y C. hirta (L.) D. Don. (hierba de unos 80cm) en la selva de Dois Irm![]() os, municipio de Recife, restos de pluvisilva oceánica en el estado de Pernambuco ( 8°00'00''-8°7'30'' S; 34°52'30' W). Las observaciones de campo se realizaron entre los meses de enero de 1993 y enero de 1995.
os, municipio de Recife, restos de pluvisilva oceánica en el estado de Pernambuco ( 8°00'00''-8°7'30'' S; 34°52'30' W). Las observaciones de campo se realizaron entre los meses de enero de 1993 y enero de 1995.
Seleccionamos diez individuos de cada especie para el registro quincenal de las fenofases de floración y fructificación. Hemos considerado como periodo de floración, el intervalo de tiempo entre el comienzo del desarrollo de los botones, hasta la ausencia completa de flores en antesis; y el periodo de fructificación, desde el comienzo del desarrollo de los frutos hasta la madurez total de los mismos.
Las observaciones sobre el horario y la duración de la flor se realizaron a partir de flores en pre-antesis, marcadas y observadas hasta el inicio de la marchitez, en cinco flores de los diez individuos. La viabilidad polínica se obtuvo mediante el test de teñibilidad en carmín acético al 2% (Radford et al. 1974). El cociente polen/óvulo (P/O) y el índice de reproducción cruzada se hallaron según las técnicas de Cruden (1977). Las anteras de viente flores separadamente fueron abiertas y su contenido vaciado en una solución (3: 1, v/v) de ácido lático y glicerina de acuerdo con Lloyd (1972) e inmediatamente se registró el número de granos de polen (Maeda 1985), con un hemacitómetro (Cámara de Neubauer).
Para el estudio del sistema reproductor se realizaron experimentos de polinización controlada (auto-polinización espontánea y manual y polinización cruzada), donde los granos de polen fueron retirados de las anteras y depositados sobre el estigma receptivo. La existencia o no de agamospermia fue comprobada siguiendo a Radford et al. (1974). En todos los experimentos las flores fueron protegidas con bolsas de papel semipermeable hasta un dia después de la antesis.
Los análisis meyóticos se hicieron fijando botones florales en una solución de alcohol absoluto/ácido acético glacial/carmín acético (10: 4: 1, v/v/v) añadiendo una gota de acetato férrico como mordiente. Después de 15 hr, se procedió a la tinción del material en carmín acético (2%) durante 30 min, tras la cual fue montado en láminas histológicas para su observación (Luceño & Castroviejo 1991).
Un exicata testigo de cada especie se conserva en el herbario UFP (n 08895, 09272 y 10703).
Resultados
El período de floración de las tres especies se extiende a lo largo de todo el año. En C. capitellata la intensidad de floración fue uniforme durante todo el año, mientras que en las otras dos especies hubo picos de floración: abril y julio de 1993 y abril y mayo de 1994 para C. bullosa y marzo y abril para C. hirta. Paralelamente al periodo de floración, la fructificación se extendió durante casi todo al año, con variaciones en el porcentaje de frutos según la fenofase.
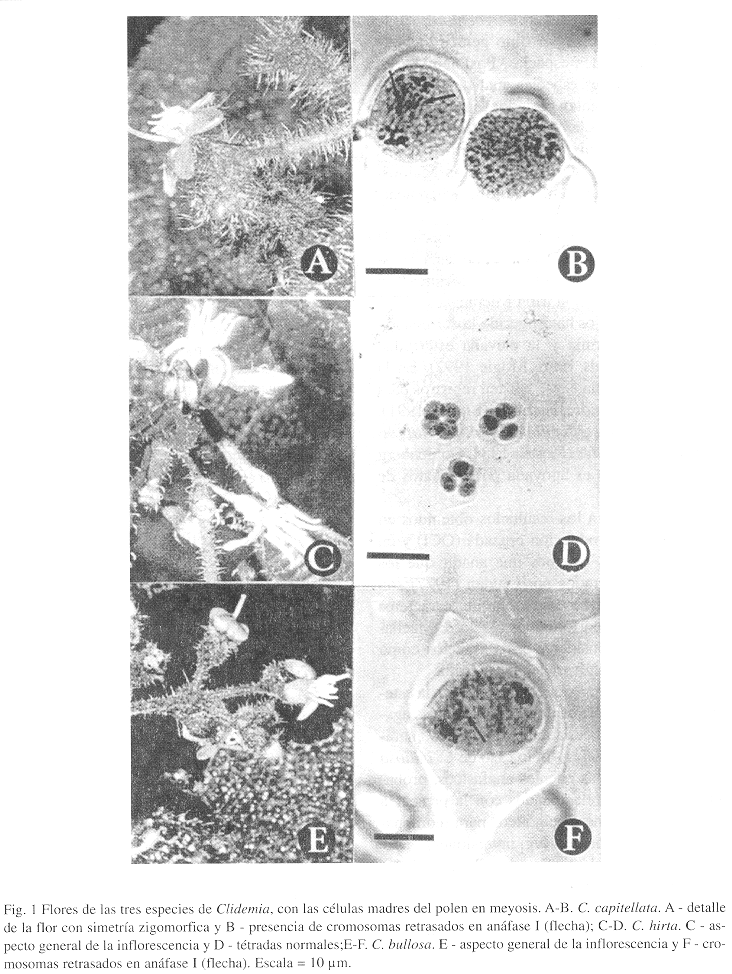
Las especies aquí estudiadas presentan grandes semejanzas en cuanto a los caracteres vegetativos, y las flores se disponen en tirsoides axilares (Figs. 1A, 1C y 1E), presentan antesis diurna (5: 00 hr) y duran más de 12 hr.
En el área estudiada, las tres especies se distribuyen por los márgenes de la selva y crecen siempre juntas, formando pequeñas poblaciones. De hecho, se registraron visitas esporádicas a las tres especies, de abejas del género Augochloropsis, que recogen polen por vibración ("buzz pollination").
Los resultados de la prueba de agamospermia demuestran que las tres especies estudiadas son agamospérmicas, no seudogámicas, ya que las porcentajes de frutos formados en el test de apomixia y en el de auto-polinización manual son semejantes.
Los resultados obtenidos en el índice de reproducción cruzada (OCI) y en el cociente P/O, que califican las especies como xenógamas.
Los estudios meióticos realizados en las tres especies muestran que en C. bullosa y C. capitellata hay cromosomas retrasados en anáfase I (Fig. 1B y 1F) con numerosas figuras cromosomáticas distintas del bivalente en metáfase I y formación de tétradas con micronúcleos, como exponente de una meiosis irregular, lo que tiene como consecuencia una elevada esterilidad cromosomática. En C. hirta la meiosis es regular (Fig. 1D), con formación de tétradas normales. En C. bullosa y C. capitellata fueron observadas varias células con puentes anafásicos.
Discusión
La distribuición de las tres especies por los márgenes de la selva las encuadra como especies simpátricas que, probablemente, comparten recursos e incluso el mismo vector polinizador (Grant 1989).
Según la revisión hecha por Renner (1989), la polinización por abejas predomina en las Melastomatáceas (98%); el 25% de las 57 especies de Miconieae estudiadas, son visitadas por abejas del género Augochloropsis, lo que podria provocar un flujo génico elevado entre las diferentes especies. Posiblemente es el caso de las tres especies analizadas en este trabajo, que presentan periodos fenológicos superpuestos y semejanza en la morfologia floral y comparten los mismo vectores polinizadores.
Las tres especies estudiadas son agamospérmicas y pertenecen a la tribu Miconieae, apoyando lo inventario de Goldenberg (1994) sobre el sistema reproductivo de especies de Melastomatáceas que encontró, de las 100 especies estudiadas, 39 son agamospérmicas (el 87,2% pertenecen a la tribu Miconieae).
Varios trabajos han sugerido la correlación entre agamospermia y la elevada esterilidad polínica (Richards 1986, Mogie 1992). En la familia Melastomataceae existen registros para las especies Leandra australis (Borges 1991), Miconia albicans, M. pohliana, M. stenotaschya y M. ligustroides (Renner 1984, Goldenberg 1994). Esta idea es apoyada por los datos de este artículo.
En relación a los resultados obtenidos en el índice de reproducción cruzada (OCI) y en el cociente P/O, tenemos que añadir que los criterios considerados por Cruden (1977) son imperfectos por basarse solo en caracteres cuantitativos y morfológicos (Melo & Machado 1998), sin considerar otros aspectos como la esterilidad polínica y agamospermia.
Otro dato citogenético que refuerza la esterilidad cromosomática de muchas especies agamospérmicas es la aparición de puentes anafásicos (Stebbins 1971). En lo caso de C. bullosa y C. capitellata las puentes anafásicos, probablemente, están relacionados con la presencia de cromosomas heterocigóticos para reordenaciones cromosomáticas (vg. inversiones).
En relación al posible origen híbrido de las especies agamospérmicas en las Melastomatáceas, no existen datos concluyentes. Los únicos registros meyóticos, fuera lo presente trabajo, son los de Goldenberg (1994) en Miconia fallax y M. stenotaschya. Este autor encontró irregularidades meyóticas en las especies y sugirió que serían producto de hibridaciones.
Resumen
Se estudió el sistema reproductivo de Clidemia capitellata (Bonpl.) D. Don, C. bullosa DC. y C. hirta (L.) D. Don. Las tres especies se distribuyen en las márgenes de la selva de Dois Irm![]() os, Recife, Pernambuco, Brasil donde forman pequeñas poblaciones y se pueden calificar como especies simpátricas. El periodo de floración se extiende todo el año. Hay visitas esporádicas de abejas Halictidae (Augochloropsis sp.) que recogen polen por vibración ("buzz pollination"). Las tres especies son agamospérmicas y muestran un elevado índice de esterilidad masculina, medida por la escasa viabilidad polínica.
os, Recife, Pernambuco, Brasil donde forman pequeñas poblaciones y se pueden calificar como especies simpátricas. El periodo de floración se extiende todo el año. Hay visitas esporádicas de abejas Halictidae (Augochloropsis sp.) que recogen polen por vibración ("buzz pollination"). Las tres especies son agamospérmicas y muestran un elevado índice de esterilidad masculina, medida por la escasa viabilidad polínica.
Referencias
Baumgratz, J.F.A. & N.M.F., Silva,. 1986/88. Ecologia da polinizaç![]() o e biologia da reproduç
o e biologia da reproduç![]() o de Miconia stenotaschya DC. (Melastomataceae). Rodriguesia 64/66 (38/40): 11-23. [ Links ]
o de Miconia stenotaschya DC. (Melastomataceae). Rodriguesia 64/66 (38/40): 11-23. [ Links ]
Borges, H.B.N. 1991. Biologia reprodutiva de quatro espécies de Melastomataceae. Dissertaç![]() o de Mestrado. Universidade Estadual de Campinas, S
o de Mestrado. Universidade Estadual de Campinas, S![]() o Paulo. [ Links ]
o Paulo. [ Links ]
Cruden, R. W. 1977. Pollen-ovule ratios: a conservative indicator of breeding systems in flowering plants. Evolution 31: 32-46. [ Links ]
Ernst, A. 1918. Bastardierung als Ursache der Apogamie im Pflanzenreich. Gustav Fischer, Jena. [ Links ]
Goldenberg, R. 1994. Estudos sobre a biologia reprodutiva de espécies de Melastomataceae de Cerrado em Itirapina . Dissertaç![]() o de Mestrado. Universidade Estadual de Campinas, S
o de Mestrado. Universidade Estadual de Campinas, S![]() o Paulo. [ Links ]
o Paulo. [ Links ]
Grant, V. 1989. Especiación Vegetal. México. Limusa, México, D.F. 578 p. [ Links ]
Gustafsson, A. 1946/47. Apomixis in higher plants. L. Univ. Arsskrift 42-43: 1-370. [ Links ]
Lloyd, D.G. 1972. Breeding systems in Cotula L. (Compositae, Anthemideae). New Phytol. 71: 1181-1194. [ Links ]
Luceño, M. & S., Castroviejo. 1991. Agmatoploidy in Carex laevigata (Cyperaceae). Fusion and fission of chromosomes as the mechanism of cytogenetic evolution in Iberian populations. Pl. Syst. Evol. 177: 149-159. [ Links ]
Maeda, J.M. 1985. Manual para uso da câmara da Neubauer para contagem de pólen de espécies florestais. Departamento de Silvicultura. Escola Superior de Agricultura "Luiz de Queiroz". Universidade de S![]() o Paulo. Piracicaba, S
o Paulo. Piracicaba, S![]() o Paulo. [ Links ]
o Paulo. [ Links ]
Melo, G.F.A. & I.C.S. Machado. 1996. Biologia da reproduç![]() o de Henriettea succosa DC. (Melastomataceae). Rev. Bras. Biol. 56(2): 383-389. [ Links ]
o de Henriettea succosa DC. (Melastomataceae). Rev. Bras. Biol. 56(2): 383-389. [ Links ]
Melo, G.F.A. & I.C.S. Machado. 1998. Auto-incompatibilidade em Miconia ciliata (L.C. Rich.) DC. (Miconieae-Melastomataceae). Acta bot. bras. 12(2): 113-120. [ Links ]
Mogie, M. 1992. The evolution of asexual reproduction in plants. Chapman & Hall Press. London. 276 p. [ Links ]
Radford, A.E., W.C., Dickison,J.R.Massey & C.R. Bell. 1974. Vascular plant systematics. Harper & Row Publishers. New York. 891p [ Links ]
Renner, S. S. 1984. Phaenologie, Bluetenbiologie und Rekombinationsysteme einiger zentralamazonischer Melastomataceen. Tesis de Dotorado. Universität Hamburg. [ Links ]
Renner, S.S. 1989. A survey of reproductive biology in neotropical Melastomataceae and Memecylaceae. Ann. Missouri Bot. Gard. 76: 496-518. [ Links ]
Renner, S.S. 1993. Phylogeny and classification of the Melastomataceae and Memecylaceae. Nord. J. Bot. 13: 519-540. [ Links ]
Richards, A.J. 1986. Plant breeding systems. Unwin & Allem. London. 529 p. [ Links ]
Sell, Y. & G. Cremers. 1987. Les inflorescences des Melastomataceae guyanaises, leur filiation et leur valeur taxonomique. Can. J. Bot. 65: 999-1010 [ Links ]
Stebbins,G.L. 1971. Chromosomal evolution in higher plants. Edward Arnold, London. 468 p. [ Links ]
Whiffin, T. 1973. Analysis of a hybrid swarm between Heterocentron elegans and H. glandulosum (Melastomataceae). Taxon 22: 413-423. [ Links ]
1 Departamento de Botânica-CCB/UFPE, Cidade Universitária. 50.372-970. Recife/PE. Brasil. Fax: 550812718348. E-mail:gfmelo@npd.ufpe.br
2 Área de Botánica. Facultad de Ciencias Experimentales, Universidad Pablo de Olavide Real. Ctra. de Utrera, Km 1 E-4100- Sevilla, España.

