











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.46 no.4 San José dic. 1998
Abstract
A lower mastodont molar was found in Nacaome river, Nicoya, Guanacaste, Costa Rica. It is a Pleistocene specimen of Cuvieronius hyodon. C. hyodon and Haplomastodon waringi, mountain and savanna mastodonts respectively, have been found in the same place. This evidence and the dental morphology suggest that they were seasonaly simpatric species with different alimentary niches. Probably H. haringi fed on savanna or prairie plants (very ligneous with high silica concentration, e.g. Cyperaceae ) whereas C. hyodon consumed softer plants (the enamel is less damaged).
Key words
Cuvieronius hyodon, Haplomastodon waringi, Gomphotheridae, Proboscidea, dental morphology, Pleistocene, Nacaome, Costa Rica.
Desde 1978 se han recolectado varios restos fósiles en la ribera y el lecho del Río Nacaome (Nicoya), Costa Rica. Entre ellos hay osamentas de un teleósteo (Laurito 1988, Laurito et al.1993 y Alvarado 1994), de un quelonio correspondiente a Rhinoclemmys nicoyana (Acuña-Mesén y Laurito 1996), de caballos del género Equus (Laurito et al. 1993) y otros, que han sido agrupados en una asociación faunística, parecida a la localizada desde 1940 en el Bajo de los Barrantes (Costa Rica). Los restos más completos de mastodonte encontrados en este río fueron identificados primero por Mora (1981) como pertenecientes a Mammuthus americanum. Más tarde, Laurito (1990) hizo una revisión y los identificó como Cuvieronius hyodon. Sin embargo, Lucas y Alvarado (1991), Alvarado (1994) y Lucas, Alvarado y Vega (1997), los identifican como Haplomastodon waringi, reportando por primera vez esta especie en América Central. Hasta la fecha no se había informado de ningún fósil de C. hyodon en el Río Nacaome. No obstante, esta especie se ha encontrado en diferentes sitios de Costa Rica (Laurito1988 y Alvarado 1994). Varias giras de inspección a lo largo del río durante 1996 y 1997, propiciaron la colecta de una pieza fósil nueva, motivo de nuestro estudio. Este tiene como finalidad describir el especímen, analizar la llegada de C. hyodon a Costa Rica y discutir sobre la coexistencia en simpatría de éste y H. waringi.
Se hizo localización visual in situ en setiembre de 1997 en la corriente del Río Nacaome, en San Juan de Nicoya, Costa Rica (10° 08 55 N; 85° 19 07 W). No se determinaron superficialmente otros restos en los alrededores. El fósil se depositó en la Escuela de Biología de la Universidad de Costa Rica ( AN-P 0001). La identificación de su contexto geológico y cronológico se hizo con base en Alvarado (1994) y Laurito (1990). Se estudió su morfometría, la morfología cualitativa (coloración, relieve, conos y cónulos), se confeccionaron varios registros fotográficos desde diferentes ángulos visuales (dorsal, anterior, posterior y lateral) y se realizó una comparación anatómica respecto a H. waringi por encontrarse en el mismo contexto.
Contexto geológico y cronológico: En el Río Nacaome aflora un conglomerado fluvial del Cuaternario con sedimentos del Pleistoceno Superior ( Mora 1981, Alvarado 1994 y Laurito et al. 1993), período en que probablemente vivió el individuo al que pertenece el fósil.
Ubicación taxonómica:Proboscidea: Gomphotheriidae: Cuvieronius hyodon.
Ubicación anatómica y características cuantitativas y cualitativas: Es la corona del molar inferior izquierdo (1M), rescatado en sus dos terceras partes. Simpson y Couto (1957) recalcan que aisladamente, los dientes de Cuvieronius y Haplomastodon son difíciles de diferenciar. A pesar de ello, los dos géneros pueden ser distinguidos primordialmente por sus defensas. Las de Cuvieronius poseen bandas de esmalte que forman una espiral mientras que las de Haplomastodon son rectas en los juveniles o ausentes en los adultos. Para Lucas, Alvarado y Vega (1997), el asignar la mayoría de los proboscídeos costarricenses a C. hyodon es una decisión conservadora, porque muchos de los especímenes (pero no las defensas) podrían pertenecer a Haplomastodon waringi. En Costa Rica se localizó un ejemplar de esta especie, la cual es rara en América Central. Diferenciar ambas especies, basándonos solo en los molares tiene limitaciones. Sin embargo, las piezas presentan rasgos que favorecen la realización de este trabajo (número de lofos, morfometría, características esmálticas, grado de desgaste, etc). Por ejemplo, los molares de Cuvieronius son bunolofodontes o trilofodontes, mientras que los de Haplomastodon pueden ser adicionalmente tetralofodontes o pentalofodontes (Correal 1981). Nuestro especimen presenta trece elementos cónicos (Fig.1 A-E). En el Cuadro 1, se indican las determinaciones morfométricas del especimen colectado:
| Longitud (mm) | |
| Ancho máximo | |
| Ancho mínimo | |
| Altura máxima de las cúspides | |
| Altura mínima de las cúspides | |
| Longitud anteroposterior máxima | |
| Longitud anteroposyterior mínima | |
- * Se basan en las 2/3 partes del molar.
Según Correal (1981), también Cuvieronius se identifica porque sus molares tienen series dobles de conos opuestos. Especialmente en la maxila los internos son más fuertes y tienen cónulos accesorios. En la mandíbula los más fuertes son los externos, los cuales también poseen cónulos accesorios ( Fig. 1 A-E). El molar en estudio se caracteriza porque la superficie de los conos y cónulos presenta poco desgaste en el esmalte, manifestado en las numerosas curvas del relieve que recorren el ejemplar, excepto en las cúspides (Fig.1 D). Este rasgo es determinante para identificar el fósil como perteneciente a C. hyodon y no a H. waringi, lo cual está de acuerdo con lo indicado por Correal (1981). Otras características muy útiles, que permiten diferenciar C. hyodon y H.waringi aparecen a continuación:
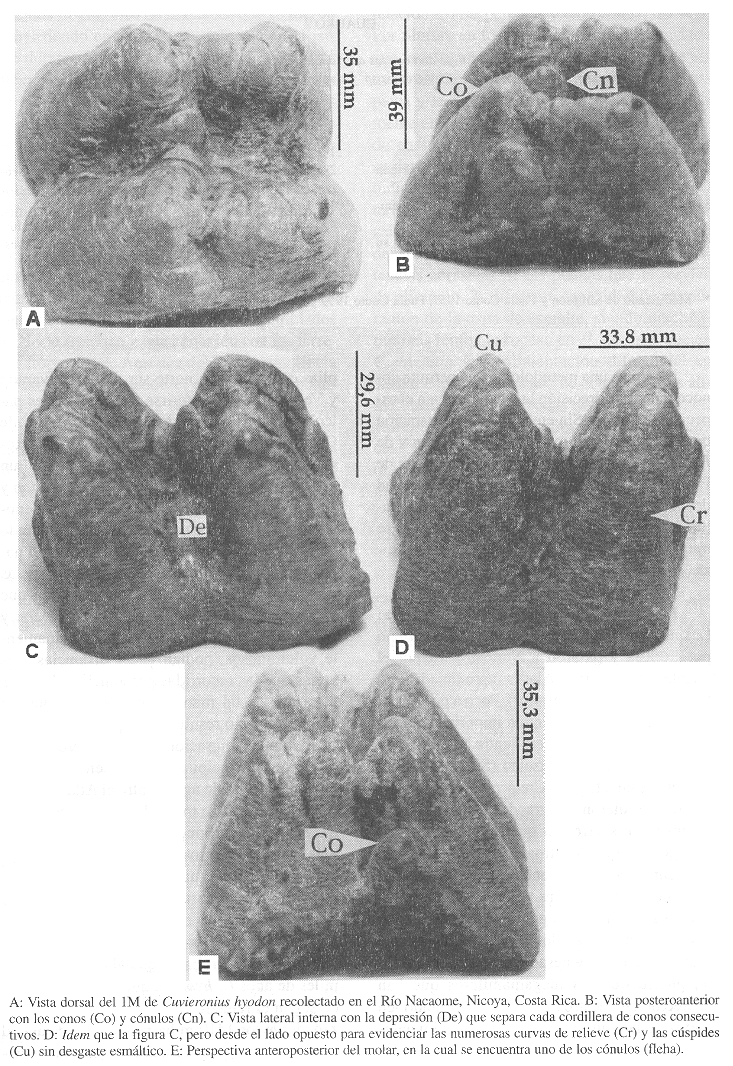
A: Vista dorsal del 1M de Cuvieronius hyodon recolectado en el Río Nacaome, Nicoya, Costa Rica
B: Vista posteroanterior con los conos (Co) y cónulos (Cn). C: Vista lateral interna con la depresión (De) que separa cada cordillera de conos consectivos. D: Idem que la figura C, pero desde el lado opuesto para evidenciar las numerosas curvas de relieve (Cr) y las cúpides (Cu) sin desgaste esmáltico. E: Perspectiva anteroposterior del molar, en la cual se encuentra uno de los cónulos (flecha).
| | |
| 1-Cráneo deprimido. | 1-Cráneo elevado tipo elefantoide |
| 2-Sínfisis mandibular corta. Incisivos superiores con banda de esmalte en toda su longitud | 2-Sínfisis mandibular corta. Incisivos superiores con sección oval. Incisivos inferiores ausentes. |
| 3- Incisivos ligeramente encorvadosa hacia abajo en ejemplares juveniles. | 3- Incisivos casi derechos en los juveniles. |
| 4- Incisivos divergentes en adultos, retorcidos sobre su eje. Banda de esmalte en espiral. | 4- Incisivos curvados hacia arriba en los adultos, sin torción helicoidal. Incisivos con estrecha banda de esmalte en los jóvenes. Sin esmalte en adultos. |
No existe una metodología que permita conocer con toda precisión la edad que tenía el especimen al morir. Sin embargo, la comparación que realizamos con molares de C. hyodon y de H. waringi, localizados en los Laboratorios de Paleontología y Arqueología de la Universidad de Costa Rica y otros que aparecen en la literatura consultada, nos indica claramente que se trata de un especímen adulto muy grande y quizás de más edad que aquellos.
La llegada de C. hyodon a Costa Rica: Los mastodontes migraron durante el Mioceno por Eurasia y pasaron a Norteamérica por el puente entre Siberia y Alaska (aproximadamente desde el Mioceno Inferior). Poco a poco Centro América fue adquiriendo importancia en la dispersión de los mamíferos entre Norte y Suramérica durante el Cenozoico (Simpson 1965). Fue precisamente en el Cretáceo-Paleoceno tardío que se dieron las grandes migraciones de los mamíferos terrestres que ocuparon el archipiélago volcánico Centroamericano, el cual estaba aún rodeado por anchas brechas marinas (Alvarado 1994 y Lucas y Alvarado 1994). Dicho procedimiento migratorio fue motivo de discusión en el pasado por considerarse que eran animales muy pesados. Esto es importante, puesto que los megamamíferos que han aparecido en nuestro istmo son el resultado -en parte- de las migraciones incipientes iniciales pero sobre todo de otras posteriores más amplias con dirección norte-sur. Lucas, Alvarado y Vega (1997), al referirse a las afinidades paleobiogeográficas de los megamamíferos de Costa Rica establecen que muchos de los táxones pleistocénicos importantes, tuvieron un origen norteamericano, entre ellos C. hyodon y H. waringi. Los proboscídeos de tallas menores, que tenían un volumen grande comparado con el de otros mamíferos, podían cruzar a nado los pasadizos marinos localizados entre las islas. Aún más, Johnson (1978), explica que inclusive los mastodontes (como C. hyodon y H. waringi) y los elefantes, que son sumamente voluminosos, podían nadar distancias más largas que las recorridas por aquellos de tallas menores, de tal manera que, la condición de archipiélago no resultó un impedimento significativo a las migraciones que se dieron en esta región a principios del Plioceno. El cierre del intercambio de aguas entre el Atlántico y el Pacífico ocurrido a mediados del Plioceno (entre 3.5 a 3.1 millones de años), facilitó aún más los movimientos migratorios. Durante este período, ocurrió una de las migraciones masivas de más importancia a través del istmo. Consideramos que esta fue la época en que tanto C. hyodon como H. waringi llegaron a Costa Rica. Es probable que al cabo de varios miles de años C. Hyodon, que era más dinámico según el registro fósil- pues se le ha localizado desde los 1410 m hasta los 10 m de altitud (Alvarado 1994)- presentara localmente movimientos migratorios de manadas, que se fueron convirtiendo en costumbres internas en el istmo durante el Pleistoceno. En este grupo posiblemente las migraciones locales se originaron en las alturas medias de nuestro país, como las faldas de las cordilleras y el Valle Central que en ese entonces estaban cubiertos de bosques (de ahí su nombre de mastodonte de las montañas), pasando luego hacia las partes bajas, como las llanuras del Pacífico y del Atlántico.
¨ Existió la probabilidad de simpatría entre C. hyodon y H. waringi? La asociación faunística encontrada en el Río Nacaome, sugiere continuidad con los territorios templados de Norte América y una conexión con los terrenos tropicales de América del Sur. La presencia de caballos y mastodontes no permite concluir que el clima tendiera a la aridez. Sin embargo, se puede suponer que el desarrollo de sabanas para la vertiente pacífica (en la región aledaña al Río Tempisque y Golfo de Nicoya, en donde desemboca el Nacaome), fuera mayor que en la actualidad. Dicho escenario permitió la pastura de manadas de ungulados y proboscídeos (Gómez 1986 y Laurito et al. 1993). Debido a esta circunstancia, en esta región pudo darse ocasionalmente la interacción simpátrica. Existen varias razones para pensar que fue así: a- los hallazgos fósiles apoyan esta idea, b- los rasgos externos del deterioro de la dentición de H. waringi indica que este ocupaba un nicho alimentario distinto, lo cual era ventajoso como se explica adelante, c- la simpatría no era permanente sino que se producía estacionalmente reduciendo las interacciones entre ambos grupos, d- Centro América muestra abundancia de proboscídeos fósiles: Mammuthus en Honduras, El Salvador, Nicaragua y Costa Rica; Stegomastodon en Nicaragua; Serridentinus en Guatemala, Haplomastodon en Costa Rica y Cuvieronius en Guatemala, Honduras, El Salvador, Nicaragua y Costa Rica. La presencia de este tipo de formas fósiles en un istmo tan estrecho y en un extensión tan reducida, unido a la gran capacidad migratoria latitudinal de C. hyodon, también sugiere una gran probabilidad de simpatría de esta especie con relación a otras pero sobre todo con H. waringi. Por último, en todos los molares de C. hyodon que investigamos es evidente que la capa de esmalte está muy bien conservada y es muy conspicua y la de Haplomastodon muestra normalmente un deterioro precoz muy notable, reflejado en su adelgazamiento o ausencia. Esto nos sugiere cuatro posibilidades: 1- que el molar estudiado se encontraba en una posición muy posterior en la mandíbula y por lo tanto no se utilizaba tanto, 2- que existía una menor resistencia del aparato masticatorio de H. waringi a las sustancias abrasivas de las plantas que le servían de alimento, 3- diferente capacidad de restauración de la capa de esmalte, siendo muchísimo más lento el proceso en H. waringi que en C. hyodon y 4- alimentación diferencial en ambas especies derivada de ocupar nichos alimentarios distintos. Todas las alternativas, las vemos como factibles. Sin embargo, en todo caso, la preservación esmáltica sigue siendo una característica peculiar de C. hyodon. Como condición sinérgica,éste probablemente consumía plantas con menos sílice o con menor grado de leñosidad que H. waringi. Este último, habitaba preferencialmente las sabanas o praderas, donde eran abundantes las ciperáceas que contienen una alta concentración de sílice en sus hojas. En la actualidad el ganado vacuno evita comerlas, los equinos no, causándoles un mayor desgaste dentario que a las vacas (Gómez, J. comunicación personal 1998). Lo mismo pudo suceder con los proboscídeos. C. hyodon prefería las montañas boscosas durante gran parte del año y ahí este tipo de plantas no era tan abundante. Esta posibilidad la consideramos la más razonable, porque los fósiles vegetales confirman que en el período Cuaternario había abundantes sabanas, entre las cuales además de algunos árboles como los que hay hoy en día, era posible encontrar muchas gramíneas, ciperáceas y otras plantas herbáceas que podían servir de alimento a estos megamamíferos. Como lo consideran Janzen y Martin (1982) y Janzen (1991), para complementar su dieta muchos de estos mamíferos también pudieron alimentarse de frutos funcionando como dispersadores de los mismos. Por otra parte, en las zonas bajas existe abundancia de frutos y semillas con cubiertas duras que proporcionaban resistencia a la masticación. Durante el Pleistoceno era posible encontrar dentro de las sabanas parches boscosos en los cuales forrageaba a lo largo del año y con mayor frecuencia H. waringi y menos C. hyodon, lo que también pudo traerle mucho mayor desgaste molar al primero por estar más en contacto con este tipo de frutos y semillas.
Agradecimientos
Agradecemos a Rafael Reyes Briceño y José Reyes Briceño por la recolección del especimen, a Teresita Aguilar (Geología, Universidad de Costa Rica U.C.R) y a Sergio Chávez (Arqueología, U.C.R.) por facilitarnos el examen de los fósiles y a Faustino Desinach por las fotografìas.
Referencias
Acuña-Mesén, R. A. & C.A. Laurito. 1996. Nueva especie de Rhinoclemmys Fitzinger,1836 (Chelonii, Cryptodira) del Cenozoico tardío de Costa Rica. Ameghiniana 33: 271-278. [ Links ]
Alvarado, G.E. 1988. Centroamérica y Las Antillas: puente, barrera y filtro biológico entre Norte y Sudamérica (Cretácico al Presente). Geoistmo II: 9 - 25. [ Links ]
Alvarado, G.E. 1994. Historia Natural Antigua: los intercambios biológicos interamericanos. Tecnológica de Costa Rica. Cartago, Costa Rica. 234 p. [ Links ]
Correal,G. 1981. Evidencias culturales y megafauna pleistocénica en Colombia. Fundación para la Investigación Arqueológica Nacional N° 12, Bogotá, Colombia.148 p.
Gómez, L.D. 1986. Vegetación de Costa Rica: Apuntes para una biogeografía costarricense. In L.D. Gómez (ed). Vegetación y Clima de Costa Rica.. Vol. I. Universidad Estatal a Distancia, San José, Costa Rica. 327 p. [ Links ]
Janzen, D.H. 1991. Historia Natural de Costa Rica. Universidad de Costa Rica, San José, Costa Rica. 525 p. [ Links ]
Janzen, D.H. & P.S. Martin. 1982. Neotropical anachronisms: the fruit of Gomphoteres ate. Science 215: 19 - 27. [ Links ]
Johnson, D.L. 1978. The origin of island mammals and Quaternary land bridge history of the northern channel islands, California. Quat. Res. 10: 204 - 225. [ Links ]
Laurito, C.A. 1988. Los proboscídeos fósiles de Costa Rica y su contexto en la América Central. Vínculos 14: 28 - 58. [ Links ]
Laurito, C.A. 1990. Estudio de un ejemplar de Cuvieronius hyodon hallado en el lecho del Río Nacaome, Guanacaste,Costa Rica. Rev. Geol. Amér.,Central 11: 41 - 50. [ Links ]
Laurito, C.A., W. Valerio & E. Vega. 1993. Nuevos hallazgos paleovertebradológicos en la Península de Nicoya: implicaciones paleoambientales y culturales de la fauna de Nacaome. Rev. Geol. Amér. Central 16: 113 - 115. [ Links ]
Lucas, S.G. & G.E. Alvarado. 1991. Comentarios sobre la clasificación de mastodontes de Barra Honda (=Río Nacaome), Guanacaste, Costa Rica. Rev. Geol. Amér.Central 13: 97 - 98. [ Links ]
Lucas, S.G. & G.E. Alvarado. 1994. The role of Central America in land vertebrate dispersal during the late Cretaceous and Cenozoic. Profil 7: 401 - 412. [ Links ]
Lucas, S.G., G.E. Alvarado & E. Vega. 1997. The Pleistocene Mammals of Costa Rica. Journal of Vertebrate Paleontology 17: 413 - 427. [ Links ]
Mora, S. 1981. Barrahonda. Universidad Estatal a Distancia, San José, Costa Rica. 96 p. [ Links ]
Paula Couto, C. De. 1979. Tratado de paleomastozoología. Academia Brasileira de Ciencias. Río Janeiro, Brazil. 590 p. [ Links ]
Segura, A. 1942. Sección de Geología: informe rendido a la Secretaría de Educación Pública sobre la labor realizada en 1940. Serie de Historia del Museo Nacional 1: 15 - 50. [ Links ]
Simpson, G.G. 1965. The geography of Evolution. Capricorn, Nueva York. 249 p. [ Links ]
Simpson, G.G. & C. de Paula Cuoto. 1957. The mastodonts of Brazil. Am. Mus. Nat. Hist. 112: 125 - 190. [ Links ]
1 Sección de Anatomía Comparada, Escuela de Biología, Universidad de Costa Rica, 2060, San José, Costa Rica. Fax (506)2074216, racuña@cariari.ucr.ac.cr 2 Sección de Ciencias Biológicas, Escuela Estudios Generales, Universidad de Costa Rica, 2060. San José, Costa Rica.

