










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.46 n.3 San José Sep. 1998
Abstract
The reproduction of the terrestrial snails Polymita muscarum and Polymita venusta was studied under laboratory and natural living conditions. Eight natural populations were sampled in Holguín Province, Cuba, from September 1985 through May 1990. In the laboratory, 46 P. muscarum and 25 P. venusta were kept in 32.3 cm x 22.8 cm x 30.7 cm terraria and fed fungi-infested branches. The reproduction season was July-December in the laboratory and July-January in nature. Times between copulation and oviposition were 18 days for P. muscarun and 20 days for P. venusta and for incubation, 8 days. Mean number of eggs per nest: P. muscarum 84.12, P. venusta 142.2; natality rates: 52.65 % and 19.9 % respectively.
Keys words
Polymita sp., land snail, reproduction, eggs, copulation, oviposition, behaviour.
La demanda creciente del uso de las polimitas en la confección de objetos artesanales y el estado actual de deterioro de sus poblaciones naturales, son las premisas fundamentales para el desarrollo de su cría intensiva. Esto nos permitirá hacer un uso racional de este recurso natural que propiciará la conservación de esta valiosa especie.
El conocimiento sobre los aspectos de la biología de las especies que serán objeto de cría son elementos muy importantes y por tanto puntos a priorizar en las investigaciones. Es por ello que en el Instituto de Ecología y Sistemática del Ministerio de Ciencia, Tecnología y Medio Ambiente se lleva a cabo un estudio sobre estos aspectos en P. muscarum Lea 1834 y P. venusta Gmelin 1792.
Varios estudios han tratado aspectos de la reproducción del género Polymita. Con relación al período reproductivo se ha encontrado que comprende los meses de septiembre hasta febrero (Díaz-Piferrer 1962, Valdés et al. 1986, Bidart et al. 1989) y la mayor actividad de cópula para P. pictanigrolimbata es septiembre y octubre (Bidart et al. 1989). En trabajos realizados en el campo se encontró que P. muscarum pone sus huevos en el suelo, en tocones y hendiduras de árboles vivos (Díaz-Piferrer 1962, Bidart et.al.1991). Feijoó (1984) registró de 40 a 50 huevos y Valdés et al. (1986) señaló que el promedio de huevos de P.p. roselimbata es de 34 y que el tiempo de incubación es de12 días. No existen antecedentes de estudios sobre reproducción de P. venusta.
El presente trabajo tiene el objetivo de presentar los resultados relacionados con parámetros reproductivos tales como época de reproducción, descripción de la cópula y ovoposición, tamaño de la nidada, coeficiente de eclosión y tasa de natalidad.
Materiales y Métodos
El presente estudio está basado en trabajos realizados en ocho poblaciones naturales de la provincia de Holguín y en condiciones de laboratorio, de septiembre de 1985 a mayo de 1990.
En las poblaciones naturales estudiadas se presentan diferentes formaciones vegetales tales como: Bosque semicaducifolio (El Yayal y Las Calabazas), Bosque micrófilo costero y subcostero (Gibara, Pesquero, Playa Blanca, Jururú y Los Cocos) y Bosque Xeromorfo costero (Caletones).
Para el estudio en el laboratorio se colectaron 46 ejemplares de P. muscarum en localidad "El Yayal" y 25 ejemplares de P. venusta en Cupey, ambas de la provincia de Holguín. Estas polimitas fueron colocadas en terrarios (peceras) de 32.3 cm x 22.8 cm x 30.7 cm de capacidad. El fondo se cubrió con una capa de tierra aproximadamente de 4 cm de espesor. El alimento suministrado consistió en ramas de árboles infestadas de hongos.
La limpieza de los terrarios se realizaba todas las semanas y a la vez se renovaba el alimento, diariamente se les rociaba agua para mantener alta la humedad relativa.
Para el estudio de la reproducción se tuvieron en cuenta los siguientes parámetros propuestos por Daguzan (1981):
2- Coeficiente de eclosión: Número de juveniles que eclosionan.
3- Tasa de natalidad: Número de juveniles que eclosionan / Número de huevos puestos.
4- Duración de la incubación (días): Tiempo entre la puesta y la eclosión.
5- Duración de la eclosión (días): Tiempo desde que comienza la eclosión de huevos de un mismo nido hasta que termina.
Epoca de reproducción: La época de reproducción de P. muscarum y P. venusta, en el labororatorio, comenzó en julio y se extendió hasta diciembre observándose un pico de apareamiento en octubre (Fig. 1).
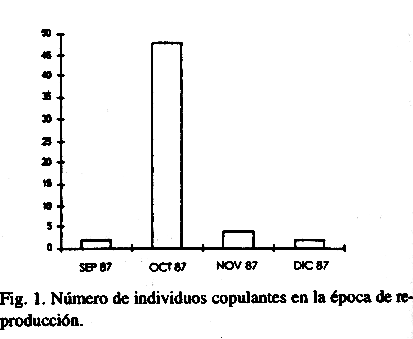
Teniendo en cuenta la relación adultos-juveniles (Cuadro 1) en las poblaciones naturales (P. muscarum) de la provincia de Holguín, podemos concluir que la época reproductiva comienza en julio con un pico en septiembre y concluye en enero, debido a la presencia de juveniles pequeños (2mm-4mm) en octubre, noviembre y diciembre. En los últimos meses del año 1985 existió un alto porcentaje de juveniles, ocurriendo lo contrario desde 1986 hasta octubre de 1987, excepto en las poblaciones de El Yayal, Gibara y Las Calabazas entre marzo y mayo de 1986 y 1987
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
| | | | | | |
Cópula: Las cópulas de P. muscarum y P. venusta se observaron con más frecuencia en octubre, sobre las paredes de los terrarios. La conducta copulatoria de P. muscarum y P. venusta es fácilmente reconocible, por la dilatación del poro genital que presentan los individuos que entran en cópula, observándose éste como un punto redondeado de color blanco que aumenta de tamaño paulatinamente.
La frecuencia de cópula por individuo es de más de una vez por época. La cópula de P. muscarum (15 observaciones) está precedida de un cortejo, el cuál se manifiesta por la gran actividad que despliegan los individuos a través del desplazamiento de uno frente al otro, en todas direcciones; movimiento de los tentáculos, rozamiento de tentáculos entre individuos, rozamiento de cuerpos, acercamiento, separación, numerosos movimientos circulares y contacto bucal. La duración media del cortejo es de 30 minutos (N =15; S = 0.96).
Una vez que ha comenzado el acercamiento, el pene de uno de los individuos se proyecta de inmediato y a medida que transcurre el tiempo, el pene aumenta de tamaño. Se ha observado en pocos individuos (4) la ausencia de movimientos circulares y preparación previa para la cópula, coincidiendo en aquellos que realizan desplazamientos verticales, sin embargo, en este caso no se mantienen todo el tiempo inmóviles, sino que por el contrario siguen un desplazamiento vertical a lo largo del eje de la planta buscando altura, con posterioridad invierten el recorrido, ambos retroceden y realizan un movimiento incompleto en sentido de las manecillas del reloj.
Una vez concluido el cortejo, se detienen los movimientos y quedan ubicados ambos individuos de forma opuesta, es decir, una con el ápice de la concha hacia arriba y la otra con el ápice hacia abajo. El pene de una de ellas se alarga primero y después el de la otra. Se produce mutuo contacto y una completa acomodación, donde el pene de cada una se inserta en la vagina de la otra, transfiriendo un espermatóforo. Las polimitas permanecen inmóviles haciendo sólo pequeños movimientos de los tentáculos.
Aproximadamente después de transcurridos 20 minutos, las polimitas comienzan a moverse para separarse, pero continúan los penes entrelazados en forma de cruceta. Después comienzan a desplazarse con movimientos circulares y poco a poco el pene va disminuyendo de tamaño y el poro genital adopta su forma habitual.
Ovoposición: El tiempo entre la cópula y la ovoposición es de 18 días (N = 15; S= 0.75) para Polymita muscarum y 20 días para P. venusta ( N= 5 ; S=1.14 ). Los desoves en el laboratorio se realizan en la tierra (sustrato)
En el laboratorio se observaron ocho desoves. Las polimitas realizan una oquedad de aproximadamente 2 cm de profundidad en el sustrato, con la parte posterior del pie al inicio y terminan con la cabeza. Después colocan el poro genital sobre la oquedad y con el cuerpo totalmente estirado e inmóvil comienzan la puesta de los huevos, uno a uno y de forma continua. Una vez depositado un grupo, hacen una pausa y después continúa el desove. Al terminar la puesta, las polimitas se alejan en busca de un sitio donde permanecen inactivas por un tiempo (1 día). No se ha observado ningún indicio de cuidado de los parentales hacia la futura descendencia.
Los huevos son redondeados, de color blanco con un diametro promedio de 2.92 mm y el peso es de 0.01g .
En una de las poblaciones estudiadas ( Los Cocos) se encontraron juveniles recien nacidos entre peciolos de Copernicia sp., acompañados de los restos de cascaras de huevos, lo que evidencia que las puestas fueron realizadas en estas plantas. Al parecer este es el sitio de puesta que habitualmente utilizan las polimitas en esta localidad, ya que la vegetación está fundamentalmente sobre roca caliza, lo que imposibilita a las polimitas realizar oquedades en el suelo y están obligadas a buscar otro sustrato que les brinde humedad y protección contra los depredadores.
Tamaño de la nidada: En el laboratorio el número de huevos por nido de P. muscarum (Cuadro 2) varió entre 32 y 172 (N = 8; X= 78; S= 7.30) y en P. venusta (Cuadro 3 ) de 95 a 182 (N=5, S=33.61).
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | ||
| | | | |
Como se puede apreciar el número de huevos por nido es muy variable. Las nidadas menores de P. muscarum (32 huevos) coinciden con los parentales más pequeños (15 mm).
La variación del número total de huevos de ambas especies, en la época reproductiva se presenta en la Fig. 4. La mayor cantidad de huevos de P. muscarum se encontró en agosto y disminuyó a medida que transcurre la época reproductiva (Fig. 2). En P. venusta fue mayor en septiembre aunque el número de huevos se mantiene más o menos estable en todo el período.
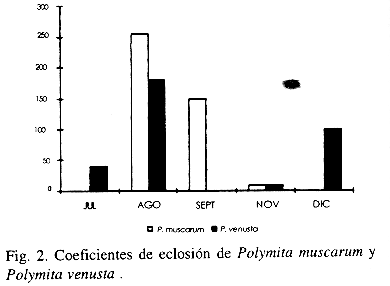
Coeficiente de eclosión y tasa de natalidad: La eclosión de los huevos en P. muscarum ocurre ocho días promedio después de las puestas (N= 8; S= 0.75). Se ha observado que la eclosión de los huevos de un mismo nido no es simultánea. La duración promedio de la eclosión de un nido es de dos días (N= 8; S=0.74) en ambas especies.
En los Cuadros 2 y 3 se presentan el coefiente de eclosión y la tasa de natalidad de P. muscarum y P. venusta respectivamente. El coeficiente medio de eclosión de P. muscarum fue de 49.12 (N=8; S=37.47) y el de P. venusta de 32. Las tasas medias de natalidad de P. muscarum y P. venusta fueron de 52.65% (N=8; S=12.77) y 19.90% (N=5; S=12.92) respectivamente.
El coeficiente de eclosión (Fig. 3) de P. muscarum disminuye en el transcurso de la época reproductiva al igual que la fecundidad. En P. venusta hay fluctuaciones de la eclosión, observándose que al inicio de la época el coeficiente es bajo, en septiembre, hay un pico que posteriormente disminuye, aumentando nuevamente en diciembre. En este caso, no hay una relación con la fecundidad, la cual permanece más o menos estable durante toda la época. El Cuadro 4 resume los parámetros estudiados en ambas especies. Como se puede apreciar en los tres primeros parámetros se observan diferencias entre las dos especies, siendo la fecundidad de P. venusta superior a la de P. muscarum, sin embargo, el coeficiente de eclosión y la tasa de natalidad son más bajos.
| | | |
| | | |
| | | |
| | | |
| | | |
| | | |
| | | |
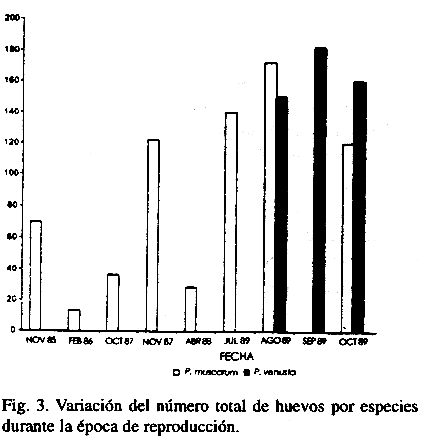
Los restantes parámetros (4, 5 y 6) son similares, sólo hay pequeñas diferencias en el tiempo entre la cópula y la ovoposición y en el tiempo de incubación.
Discusión
Epoca de reproducción: Las diferencias anuales y locales en el porcentaje de juveniles parecen deberse a que en estos meses predominan los juveniles adelantados (Bidart et.al. 1992 c) y a la intensa sequía que azotó a la provincia, incidiendo de forma negativa en la reproducción de P. muscarum. Los últimos meses de 1987 se caracterizaron por lluvias abundantes y bajas temperatura,s lo que favoreció la reproducción de P. muscarum en varias áreas de distribución como Pesquero, Gibara y Jururú.
Una frecuencia de cópula similar se presenta en el gasterópodo terrestre Helix aspersa, donde la mayoría de los individuos copulan dos veces en la misma época (Daguzan 1981).
Ovoposición: Un período similar cópula-ovoposición (16 días) señalan Valdés et al. (1986) para P. picta roseolimbata y los desoves en tierra del laboratorio reflejan lo que ocurre en la naturaleza (Díaz-Piferrer 1962). Bidart et al. (1992 b) encontraron en tocones de árboles numerosas puestas de P. muscarum y señalaron que esto podría estar dado por la gran sequedad de los suelos del lugar.
Con la observación mencionada en este trabajo se amplía el subnicho reproductivo de Polymita muscarum y se demuestra que, de acuerdo con las características ecológicas del lugar, esta especie desarrolla una estrategia para garantizar la supervivencia.
Tamaño de la nidada: Bidart et al. (1992 b) encontraron en poblaciones naturales de P. muscarum nidadas mayores a las encontradas por nosotros. Esta diferencia pudiera deberse a que los individuos de estas poblaciones presentaron tallas mayores. Según Oosterhoof (1977) algo similar ocurre para el molusco europeo Cepaea nemoralis, donde el tamaño del nido está positivamente correlacionado con el tamaño del progenitor.
Wolda (1963, 1965, 1967) y Wolda y Kreulen (1973) obtuvieron resultados similares para esta especie y señalaron además que el tamaño del nido depende de la edad de los individuos y de las condiciones ambientales en que se realizan las puestas. En nuestro estudio es posible que haya incidido el tamaño del parental en la variación del número de huevos. Sin embargo, esto debe comprobarse con mayor cantidad de individuos.
Wolda y Kreulen (1973) observaron también decrecimiento en el número de huevos de Cepaea nemoralis en la época reproductiva y lo atribuyen a cambios en la calidad y cantidad de alimento. Oosterhoof (1977), en la misma especie, señala que esto puede ser debido a que las últimas puestas las realizan los individuos de más lento crecimiento que producen nidadas menores. . Villalobos et al. (1995) encontraron para Succinea costarricana, que el número de huevos decrecía con la edad. De estos factores creemos que en nuestros resultados es más probable que haya incidido el segundo, pues el alimento durante todo el estudio se mantuvo constante. Sin embargo, no descartamos la posibilidad de que otras causas aún desconocidas pueden haber incidido.
Coeficiente de eclosión y tasa de natalidad: Al parecer el aumento del número de huevos forma parte de la estrategia reproductiva de la especie para garantizar la supervivencia. Los restantes parámetros (4, 5 y 6) son similares, sólo hay pequeñas diferencias en el tiempo entre la cópula y la ovoposición y en el tiempo de incubación. Resultados similares obtuvieron Valdés et al. (1986) en P. p. roseolimbata.
Agradecimientos
Agradecemos a Raisa Cáceres la confección de las figuras.
Resumen
Se estudiaron algunos aspectos de la reproducción de Polymita muscarun y Polymita venusta en condiciones de laboratorio y en poblaciones naturales de la provincia de Holguín durante el período comprendido entre septiembre de 1985 y mayo de 1990. Se encontró que la época de reproducción se extiende desde julio hasta diciembre en el laboratorio y desde julio a enero en la naturaleza. Se realiza una descripción de la conducta copulatoria y el desove. El tiempo entre cópula y oviposición es de 18 días para P.muscarum y de 20 días para P.venusta, el tiempo de incubación fue de 8 días. El número de huevos por nido para P. muscarum fue de 84.12 huevos y de 142.2 para P. venusta, con una tasa de natalidad de 52.65 % y 19.9 % respectivamente.
Refferences
Bidart, L., A. Socarrás, C. Iglesias , M. Reyes & M. Hidalgo Gato. 1992 a. Aspectos del ciclo de vida de Polymita muscarum muscarum y Polymita picta nigrolimbata en el laboratorio. Instituto de Ecología y Sistemática, La Habana, Cuba. 11 p. [ Links ]
Bidart, L., J. F.Milera, M. Osorio & E. Reynaldo. 1992 b. Datos reproductivos de Polymita muscarum. Instituto de Ecología y Sistemática, La Habana, Cuba. 8 p. [ Links ]
Bidart, L., M. Osorio & E. Reynaldo. 1992 c. Ecología de Polymita muscarum en la localidad "El Yayal", provincia de Holguín. Instituto de Ecología y Sistemática, La Habana, Cuba. 8 p. [ Links ]
Daguzan, J. 1981. Contribution a l' elevage de l' escargot "Petit-gris" Helix aspersa Muller (Mollusque: Gasteropode: Pulmone: Stylommatophore). I- Reproduction et eclosion des jeunes, en batiment et en condition thermohygometrioques controlees. Ann. Zootech. 30: 249-272. [ Links ]
Díaz-Piferrer, M. 1962. Reproduction of Polymita muscarum Lea, a Cuban tree snail. Caribb. J. Sc. 2: 59-61. [ Links ]
Oosterhoff, L. l977. Variation in growth rate as an ecological factor in the landsnail Cepaea nemoralis L. Neth. J. Zool. 27: 1-132. [ Links ]
Valdés, G., V. Berovides & J. F. Milera. l986. Ecología de Polymita picta roseolimbata. Cien. Biol. 15: 77-93. [ Links ]
Villalobos, C. , J. Monge-Nájera & Z. Barrientos. 1995. Life cycle and field abundance of the snail Succinea costaricana (Stylommatophora: Succineidae), a tropical agricultural pest Rev. Biol. Trop. 43: 181-188. [ Links ]
Wolda, H. (1963): Natural population of polymorphic landsnail Cepaea nemoralis (L.). Archs. Neth. Zool. 16: 381-471. [ Links ]
Wolda, H. 1965. The effect of drought on egg production in Cepaea nemoralis (L.). Archs. Neth. Zool. 16: 387-399. [ Links ]
Wolda, H. 1967. The effects of temperature on reproduction in some morphs of landsnail Cepaea nemoralis. Evolution 2: 117-129. [ Links ]
Wolda, H. & D. Kreulem. 1973. Ecology of some experimental populations of the landsnail Cepaea nemoralis (L.). II- Production and survival of eggs and juveniles. Neth. J. Zool. 23: 168-188. [ Links ]
1 Instituto de Ecología y Sistemática, Ministerio de Ciencia Tecnología y Medio Ambiente, Carretera de Varona km 3 1/2, Municipio Boyeros, CP 10800, Ciudad de La Habana, Cuba.
2 Instituto Superior Pedagógico "Félix Varela", Ministerio de Educación, Municipio Holguín, Holguín, Cuba.

