











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.46 no.3 San José sep. 1998
Abstract
The capacity of Modiolus capax to delay spawning and to hold mature gametes for long periods, was analyzed in sexually mature organisms subjected to cold water and food rations higher than those for maintenance. The experimental group, 200 mussels collected in August from Bahia de los Angeles, Baja California, Mexico, was subjected to 19±2 °C, temperature comparable to that in the natural environment during autumn and early winter; the daily food rations, delivered in open flow, were equivalent to 0.5-1.0% of the mussels dry soft body weight. The effectiveness of the experimental treatment was verified every month in a sample of 16 mussels; for contrast, a sample from Bahia de los Angeles was also analyzed. The assessed variables were general condition and gonadosomatic indices and for the histological study of the gonads, a maturity scale with eight stages was designed. In September the mussels from the natural environment were in the initial phase of post-spawning; by December they were in late post-spawning or rest. In the experimental group there were no spontaneous spawnings and general condition and gonadosomatic indices remained consistently higher than in the natural environment, however, by the eight-week of treatment the gonads showed evidences of incipient reabsorption. It is proposed that the natural reproductive cycle, mass spawning in summer and rest in late autumn and early winter, might be limiting the mussels capacity to retain mature gametes.
Key words
mussels, reproduction, delayed spawning, Modiolus capax.
El bivalvo Modiolus capax Conrad (1837) es un mitílido de amplia distribución geográfica en el Pacífico tropical de México (Brusca 1980). La abundancia de este mejillón en algunas localidades del Golfo de California ha promovido su uso como bioindicador de contaminación por plaguicidas y metales pesados en la costa este de Baja California (Gutiérrez Galindo et al. 1988, Da Costa Gómez y Valle Díaz 1989). También se han realizado una serie de estudios relativos a la biología y a la ecología de la especie principalmente con el fin de explorar la factibilidad de su cultivo (Bückle-Ramírez y Farfán 1988).
La información disponible sobre la biología reproductiva de Modiolus capax en Bahía de los Angeles (Garza Aguirre y Bückle Ramírez 1989a y b) y en la Bahía de la Paz, B.C.S. (Ochoa Báez 1985), indica que en la costa occidental del Golfo de California la liberación masiva de gametos es más intensa de junio a septiembre pero el reclutamiento de juveniles a las poblaciones naturales es escaso. Resultados similares fueron obtenidos por Aguirre Hinojosa y Bückle Ramírez (1992) utilizando colectores artificiales. Por otra parte, aunque la obtención de larvas en condiciones controladas es factible, fuera del período mayo-septiembre el porcentaje de organismos susceptibles de ser inducidos al desove se reduce drásticamente (Orduña Rojas y Farfán 1991).
En acuicultura dos de las alternativas más frecuentemente utilizadas para ampliar el período reproductivo natural en bivalvos, son la dilación del desove y la inducción a la gametogénesis. En el presente trabajo se reportan los resultados obtenidos al dilatar la liberación de gametos en especímenes sexualmente maduros de M. capax. La aproximación experimental fue la de mantener a los mejillones a una temperatura significativamente menor a la existente en el medio natural y suministrar raciones alimenticias superiores al metabolismo estándar del organismo a esa temperatura.
Materiales y Métodos
La población experimental fue recolectada en Bahía de los Angeles, B.C., México (29° 53' 33"N, 113° 31' 30"W). A partir de junio se inició la recolecta mensual de mejillones de 70 a 90 mm de longitud anteroposterior para seguir la evolución de la maduración sexual de la población natural mediante la inspección visual del volumen y coloración de la gónada. En agosto estas características fueron máximas, procediéndose a recolectar 250 mejillones de esa talla. Aleatoriamente se separaron 200 para conformar el lote experimental; el resto fue sacrificado para determinar la condición reproductiva inicial mediante los índices de condición y gonadosomático y el análisis histológico de las gónadas.
En el estanque experimental, un cilindro Nalgene de 200 l provisto con aereación continua y homogénea, se colocaron grupos de 50 mejillones en canastas de plástico instaladas en serie dentro del estanque. Para evitar la acumulación de metabolitos en el tanque experimental, sin extraer a los organismos, cada 24 hr se hizo el cambio de agua y limpieza del estanque con agua a baja presión. Antes y después de renovar el agua se midió la temperatura del agua y se verificó la ausencia de productos sexuales en el medio.
En este trabajo el régimen térmico utilizado fue de 19±2 °C, temperatura comparable a la que prevalece en Bahía de los Angeles, B.C. durante otoño y principios de invierno. La ración alimenticia diaria se estableció en 0.5-1.0% del peso seco del tejido blando del mejillón, ración que en equivalentes calóricos es superior al gasto energético de M. capax a esa temperatura (Rico Mora 1987). La ración alimenticia de microalgas se suministró en la modalidad de flujo continuo. Dependiendo de la disponibilidad, los mejillones fueron alimentados con la microalga Pavlova lutheri (Droop) Green o Tetraselmis suecica (Kylin) Butcher (26 ng cel-1 y 230 ng cel-1, respectivamente). El peso seco del tejido blando (carne y gónada) de los mejillones al iniciar el experimento era de 2.53±0.443 g.
La efectividad del tratamiento experimental en la prolongación de la condición reproductiva se verificó mensualmente en una muestra de 16 organismos. Como control, en fechas similares se colectó una muestra de 30 mejillones en Bahía de los Angeles. El experimento se prolongó hasta diciembre, mes en que del lote experimental se extrajo una muestra con varios especímenes macroscópicamente indiferenciados (gónada flácida y opaca).
En las muestras de campo y de laboratorio se estimó el Índice Gonadosomático (I.G.) y el Índice de Condición General (I.C.G.) de acuerdo a las relaciones de Sastry (1978) y Mann (1978), respectivamente. Para el análisis histológico de las gónadas (12 especímenes por mes) se siguió la técnica topográfica Hematoxilina de Harris-Eosina alcohólica.
Histología: Los cortes histológicos de las gónadas fueron inicialmente sometidas a un reconocimiento general determinándose ausencia o presencia de células sexuales y su estado de madurez, así como forma y talla de las mismas.
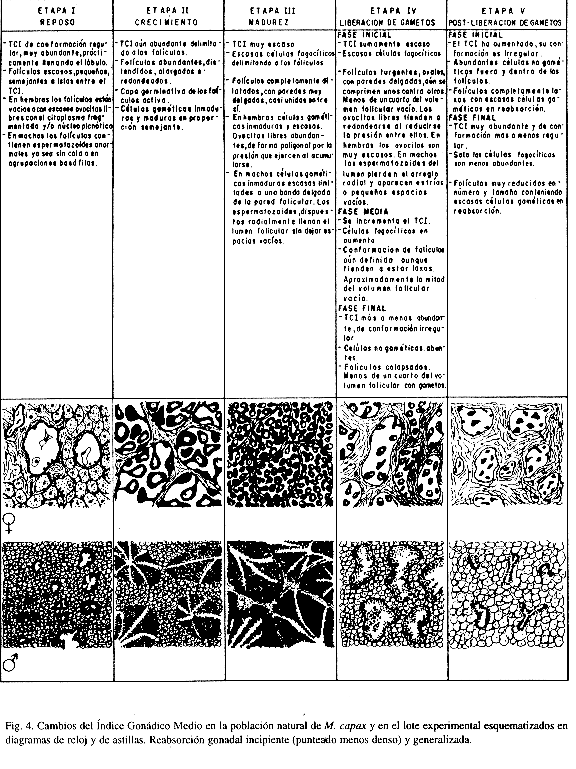
La escala de madurez utilizada en este trabajo contiene ocho estadios (cinco etapas y cuatro subetapas): Reposo, Maduración, Reproducción, Liberación de Gametos (Inicial Media y Final) y Post-Liberación de Gametos (Inicial y Media). La subdivisión de las fases de Liberación y Post-Liberación de Gametos, obedece a los objetivos del trabajo y a la época del año en que éste se llevó a cabo. La Fig. 1 contiene los esquemas y las características de las fases reproductivas utilizadas.
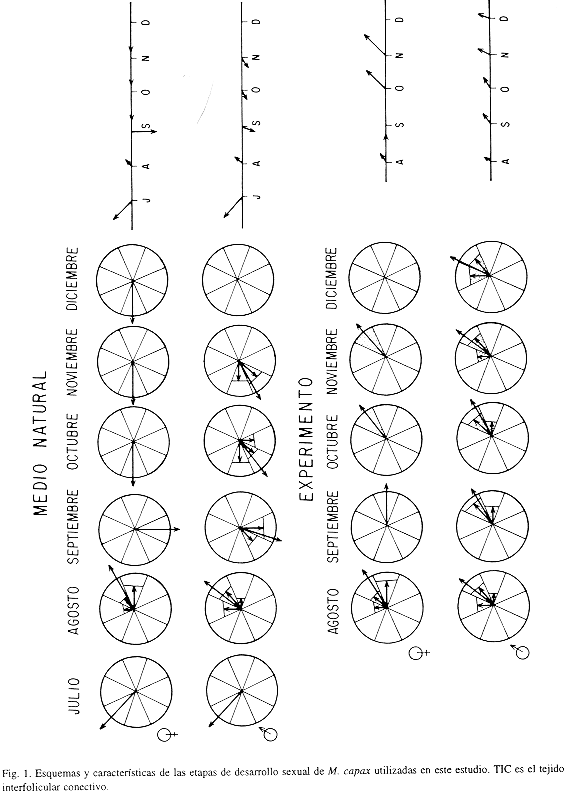
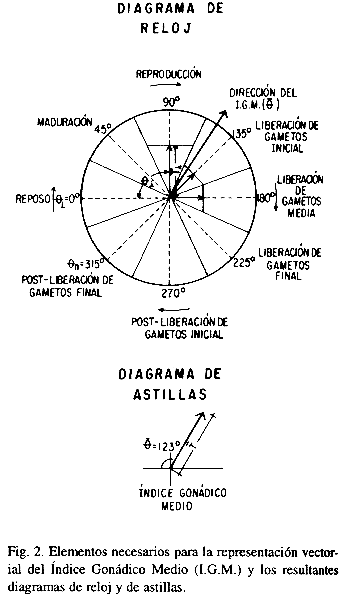
Con la información del estudio histológico se estimó un Índice Gonádico Medio (I.G.M.) basado en el método propuesto por Olguín Espinoza (1987) el cual trata las etapas reproductivas como vectores. A diferencia del ínidice utilizado por Chippefield (1953) y Seed (1969), entre otros, con este I.G.M. a la escala de madurez se le imprime magnitud y dirección, obteniéndose así una representación a manera de reloj o un diagrama de astillas. El procedimiento utilizado difiere del de Olguín Espinoza (1987) en la forma de calcular el vector medio o I.G.M.; el método propuesto es más sencillo y directo y genera resultados comparables.
El procedimiento (Fig. 2) consiste en posicionar la escala de madurez en un círculo subdividido equitativamente entre el número de etapas y subetapas (n). El límite numérico de cada estadio se hace equivalente a la posición central de la subdivisión correspondiente en grados, empezando con 0° para la primera etapa e incrementándose 360°/(n) en cada estadio. Esto es equivalente a la dirección del vector (æi) y la frecuencia relativa de organismos en ese estadio de desarrollo es la magnitud correspondiente (æi).; consecuentemente el área del sector circular es A = /(n) cuando r = 1. Para cada período de muestreo se construye un reloj con la información anterior. El vector medio constituye el I.G.M.; para el cálculo de éste en el método original primeramente se requiere estimar las coordenadas x, y. Si se utiliza un sistema de referencia cartesiano las relaciones trigonométricas a utilizar son:
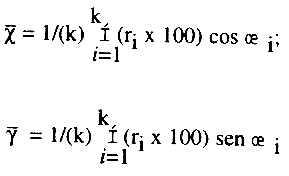
donde k es el número de etapas de desarrollo encontrado en la muestra. Por lo tanto la dirección del vector medio es ![]() magnitud es
magnitud es ![]() Esta última es una medida de dispersión del I.G.M; a mayor magnitud menor dispersión y viceversa.
Esta última es una medida de dispersión del I.G.M; a mayor magnitud menor dispersión y viceversa.
Si el sistema de referencia no es cartesiano el cálculo de ![]() se complican. Con el método propuesto la estimación del vector medio o I.G.M. se reduce a:
se complican. Con el método propuesto la estimación del vector medio o I.G.M. se reduce a:
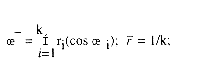
es decir la dirección del I.G.M. es simplemente la suma del producto de magnitudes por direcciones y su magnitud el recíproco del número de etapas observadas en la muestra. Esta simplificación es posible debido a que ri no es una magnitud absoluta (e.i.- número de organismos en la etapa de desarrollo en cuestión) sino una normalización de ésta (frecuencia relativa).
Resultados
El incremento en el Índice de Condición General e Índice Gonadosomático determinado en los mejillones recolectados en Bahía de los Angeles, B.C. entre junio y agosto (de 10.71 a 13.84% y de 7.31 a 38.90%, respectivamente) fue congruente con la mejora en la condición reproductiva de la población inferida de la inspección visual del volumen y coloración de la gónada efectuada en el campo (Fig. 3a-b).
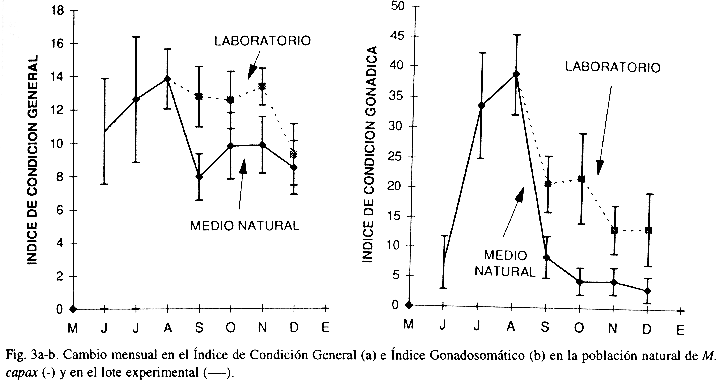
En septiembre, en los organismos del medio natural el Índice de Condición General se redujo a menos de la mitad (7.95%) y el Índice Gonadosomático decayó a 8.46%; en los meses siguientes el Índice de Condición General aumentó ligeramente pero el Índice Gonadosomático continuó decreciendo hasta alcanzar su valor mínimo (3.21%) en diciembre, último mes de muestreo. En contraste, en los organismos sometidos al tratamiento experimental el Índice de Condición General se mantuvo estable hasta noviembre, mientras que el Índice Gonadosomático aunque también disminuyó a 20.80% en el mes de septiembre, en diciembre aún era tres veces más alto que el estimado en el medio natural.
El análisis histológico de las gónadas reveló que al iniciar el experimento (agosto) los especímenes se encontraban en los estadios de Reproducción y de Liberación de Gametos inicial y media (Fig. 4). En la población de hembras, el 17% se encontraba en la etapa de Reproducción, el 33% en Liberación de Gametos inicial y el 50% en Liberación de Gametos media; en los machos, el 33% estaba en la etapa de Reproducción, el 50% en la fase inicial de Liberación de Gametos y el 17% en la fase media. A pesar de estas diferencias el Índice Gonádico Medio (I.G.M.) en ambos casos se ubicó en el cuadrante de Liberación de Gametos inicial (hembras æ-= 150°; machos æ- = 128°).
En los organismos provenientes del medio natural, en los meses siguientes ni hembras ni machos presentaron evidencias de re-inicio de gametogénesis. En septiembre el I.G.M. en ambos sexos se ubicó en la subetapa de Post-liberación de Gametos inicial. De octubre a diciembre todas las hembras analizadas estaban en la etapa de Reposo y aunque en los machos se observó mayor variabilidad, el I.G.M. reveló un avance progresivo hacia esa etapa. En los organismos sometidos al tratamiento experimental, el I.G.M. estimado en ambos sexos indica que éstos se conservaron en la subetapa de Liberación de Gametos inicial observada al iniciar el ensayo (Fig. 4). Sin embargo, en octubre ya algunas hembras presentaban procesos de reabsorción incipiente y generalizados en noviembre; en los machos algunos especímenes extraídos en octubre ya presentaban reabsorción generalizada.
Discusión
Los organismos que se recolectaron en Bahía de los Angeles de octubre a diciembre estaban en reabsorción gonadal o en reposo. Estos resultados coincien con el estudio reproductivo anual realizado por Ochoa Báez (1985) en otra poblacion de Modiolus capax del Golfo de California y con los resultados de las experiencias de inducción al desove efectuadas por Orduña Rojas y Farfán (1990).
Una posible consecuencia de que la especie entre en un proceso prolongado de reabsorción y reposo gonadal es la baja capacidad de retener gametos maduros por períodos largos. En el experimento de dilación del desove, aunque la condición general y gonadosomática de M. capax se mantuvo significativamente mejor que en el medio natural, para la octava semana de tratamiento las gónadas ya presentaban procesos incipientes de reabsorción.
Períodos similarmente cortos de retención de gametas maduras se han encontrado en poblaciones de Crassostrea gigas (Paniagua Chávez 1992) y de C. virginica (Loosanoff y Davis 1951) cultivadas o transplantadas a zonas donde la temperatura del agua no propicia el desove. En circunstancias normales las poblaciones estudiadas de estos bivalvos también desovan masivamente a fines del verano y seguidamente entran a una fase de reabsorción y reposo gonadal cuya duración es de 3 a 4 meses. En contraste, las especies que después del desove principal de verano rápidamente reinician la gametogénesis e invernan con la gónada en etapas avanzadas de desarrollo, pueden tener una alta capacidad de retener gametos por periódos prolongados. Venus (Mercenaria) mercenaria cuyo ciclo reproductivo tiene esas caracterísitcas, al ser sometida a bajas temperaturas puede retener gametos maduros hasta por 9 meses (Loosanoff y Davis 1951, Menzel 1989, Manzi et al. 1991).
La engorda y maduración gonadal de moluscos bivalvos en condiciones controladas es un proceso realtivamente costoso. Para la mayoría de las especies de interés para la acuicultura se recomiendan suministros de raciones alimenticias equivalentes al 3-7% del peso seco del tejido blando del organismo por día (Utting 1993). En contraste, para completar la maduración gonadal de organismos provenientes del medio natural o mantenerles la condición reproductiva óptima, en algunos casos pude lograrse con raciones más moderadas (0.5-3%) e incluso en inanición debido a que el proceso reproductivo es prioritario y procede a expensas de las reservas del organismo; el costo puede ser la pérdida en fecundidad y/o vigor y sobrevivencia de las larvas (Bayne 1975, Lannan et al. 1980, Muramata y Lannan 1984).
En el caso de M. capax, Mazón Suástegui (1988) reporta que la última fase de maduración puede lograrse suministrando harina de arroz o trigo en raciones diarias del.12% del peso total del organismo. En cambio, según se observó en el presente estudio, la capacidad del mitílido de retener gametas maduras es limitada debido al rápido establecimiento del proceso de reabsorción gonadal. Períodos largos de reabsorción y reposo sexual natural posiblemente también dificulten la recuperación e inducción a la gametogénesis en condiciones controladas.
Agradecimientos
Las autoras agradecen a Antonio Reséndiz su apoyo en la recolección de las muestras, a Norberto Flores Acevedo su colaboración en el cultivo de microalgas y a Modesto Ortíz por sus valiosos comentarios sobre el manuscrito.
Resumen
La capacidad de Modiolus capax de dilatar el desove y retener gametos maduros por períodos largos se estudió en organismos sexualmente maduros sometidos a condiciones de baja temperatura y raciones alimenticias superiores a las de mantenimiento. La efectividad del tratamiento se verificó mensualmente y se contrastó con la condición reproductiva de los organismos en el medio natural. En el lote experimental no se registraron desoves expontáneos y aunque los índices de condición general y gonadosomático se mantuvieron consistentemente mejores que en el medio natural, para la octava semana de tratamiento las gónadas ya presentaban evidencias de reabosorción. Se postula que las caracterísitcas del ciclo reproductivo natural, desove masivo en verano y reposo a fines de otoño y pricipios de invierno, limita la capacidad del mítilido de retener gametos maduros por períodos prolongados.
Referencias
Aguirre Hinojosa, E. & L.F. Bückle Ramírez. 1992. Análisis de la fijación del mejillón Modiolus capax (Conrad) (Bivalvia-Mytilidae) durante el desarrollo de una comunidad sobre sustratos artificiales en la Bahía de Los Angeles, B.C. Ciencias Marinas 18: 33-48. [ Links ]
Bayne, B.L. 1976. Aspects of reproduction in mollusks, p. 432-448. In M. Wiley (ed.). Estuarine Processes. Vol. 1: Uses, stress and adaptation to the estuary. Academic, Nueva York.. [ Links ]
Brusca R.C. 1980. Common intertidal invertebrates of the Gulf of California. University of Arizona, Tucson. 513 p. [ Links ]
Bückle Ramírez L.F. & B.C. Farfán. 1988. Experiencias y posibilidades del cultivo de Modiolus capax en Bahía de Los Angeles, p. 119-127. In Memorias, Encuentro regional sobre producción del mejillón. Ensenada, Baja California, México. [ Links ]
Chipperfield, P.N.J. 1953. Observations on the breeding and settlement of Mytilus edulis (L.) in british waters. J. Mar. Biol. Ass. U.K. 32: 449-476. [ Links ]
Da Costa-Gómez, C.A. & N.A. Valle Díaz. 1989. Disponibilidad biológica de metales traza en el mejillón Modiolus capax del Mar de Cortéz. Tesis de Licenciatura. Facultad de Ciencias Marinas. Universidad Autónoma de Baja California. Ensenada, Baja California, México. 77 p. [ Links ]
Garza Aguirre, M. del C. & L.F. Bückle Ramírez. 1989a. Ciclo reproductivo del mejillón Modiolus capax (Conrad, 1837) (Bivalvia, Mytilidae, Anisomyaria) en la Bahía de Los Angeles, Baja California, México. An. Inst. Cienc. del Mar y Limnol., U.N.A.M. 15: 157-170. [ Links ]
Garza Aguirre, M. del C. & L. F. Bückle Ramírez. 1989b. Estructura de tallas, reclutamiento y crecimiento del mejillón Modiolus capax (Conrad) (Bivalvia: Mytilidae) en la Bahía de Los Angeles, Baja California, México. An. Inst. Cienc. del Mar y Limnol., U.N.A.M. 16: 245-254. [ Links ]
Gutiérrez Galindo, E.A., G. Flores Muñoz & J.A. Villaescusa Celaya. 1988. Hidrocarburos clorados del Valle de Mexicali y Alto Golfo de California. Ciencias Marinas 14: 77-90. [ Links ]
Lannan, J.E., A. Robinson & W.P. Breese. 1980. Broodstock management of Crassostrea gigas. II.- Broodstocks conditioning to maximize larval survival. Aquaculture 21: 337-345. [ Links ]
Loosanoff, V.L. & M.C. Davis. 1951. Delaying spawning of Lamellibranchs by low temperature. J. Mar. Res. 10: 197-202. [ Links ]
Mann, R. 1978. A comparison of morphometric, biochemical and physiological condition in marine bivalve mollusks, p. 484-497. In J.A. Thorp & I.W. Gibbons (eds.). Energy and environmental stress in aquatic systems. DOE Symp. Ser. No. 48. [ Links ]
Manzi, J.J., N.H. Hadley & R.T. Dillon. 1991. Hard clam, Mercenaria mercenaria, broodstocks: growth of selected hatchery stocks and their reciprocal crosses. Aquaculture 94: 17-26. [ Links ]
Mazón Suastegui, J.M. 1988. Acondicionamiento y desove de cuatro especies de moluscos bivalvos alimentados con dietas artificiales. Rev. Latinoam. Acuicultura. 23: 4-13. [ Links ]
Menzel, R.W. 1989. The biology, fishery and culture of quahog clams, Mercenaria, p. 507-521. In J.J. Manzi & M. Castagna (eds.). Clam mariculture in North America. Elsevier, Amsterdam.
Muranaka M.S. & J.E. Lannan. 1984. Broodstock management of Crassostrea gigas: environmental influences on broodstock conditioning. Aquaculture 39: 217-228.
Ochoa Báez, R.I. 1985. Antecedentes sobre el ciclo de reproducción de Modiolus capax (Conrad, 1837) (Bivalvia: Mytilidae) en la Bahía de La Paz, Baja California Sur, México. Inv. Marinas CICIMAR 2: 86-103. [ Links ]
Olguín Espinoza, I. 1987. The reproductive cycle of Crassostrea virginica (Gmelin) in the Apalachicola Bay, Florida. Masters thesis. Florida State University, Tallahassee. 87 p. [ Links ]
Orduña Rojas, J. & B. C. Farfán. 1991. Induced spawning and ontogeny of Modiolus capax Conrad (Bivalvia: Mytilidae). Veliger 34: 302-308. [ Links ]
Rico Mora, R. 1987. Efecto interactivo de la temperatura y de concentración de microalgas en la fisiología alimenticia y la energía potencial para el crecimiento de Modiolus capax Conrad (Bivalvia: Mytilidae). Tesis de Maestría en Ciencias. Centro de Investigación Científica y de Educación Superior de Ensenada, B.C. México. 91 p. [ Links ]
Sastry, A. N. 1978. Physiology and ecology of reproduction in marine invertebrates, p. 279-299. In F.J. Verberg (ed.). Physiological ecology of estuarine organisms. University of South Carolina, Columbia. [ Links ]
Seed, R. 1976. Ecology, p. 13-65. In: B.L. Bayne (ed.). Marine mussels their ecology and physiology. Int. Biol. Programme X. Cambridge University, Cambridge. [ Links ]
Utting, S.D. 1993. Procedures for the maintenance and hatchery conditioning of bivalve broodstock. World Aquaculture 24: 78-82. [ Links ]
1Centro de Investigación Científica y de Educación Superior de Ensenada. Km. 107 Carret. Tij.-Eda. Ensenada, Baja California, México. Fax (61) 75 05 72, cfarfan@cicese.mx.
2Centro Regional de Investigación Pesquera El Sauzal. Apdo. Postal 1306. Ensenada, Baja California, México.

