











 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.46 no.2 San José Jun. 1998
Abstract
Cryptobiota of dead fragments of five branches in live position and five fallen pieces of the coral Acropora palmata each one of approximate 1dm3, covered by filamentous algae were extracted from the north reef crest of Isla Grande (Colombian Caribbean), in April 1991. There were three groups of organisms according to size and position (on and within the coral): 1) mobile epibenthos, mainly microcrustaceans that live among the filamentous algae 2) boring microcryptobiota, located in the layer between the epilithic organisms and the coral skeleton itself and, 3) perforating macrocryptobionts that bore and penetrate the coral skeleton. Polychaetes, sipuncu-lids, mollusks and crustaceans were most abundant in the last group. There were no differences in macrocryptobiont composition between standing dead branches and fallen fragments. There was a large variation in total biomass and type and density of macro-cryptobionts, possibly associated to stochastic factors such as placement and thickness of branches and small scale variations in recruitment.
Key words
Cryptobiota, Acropora palmata, dead coral, epibenthon, Colombian Caribb ean.
The structure and shape of reefs are the result of a dynamic system of constructive and destructive forces that constantly transform them (Schuhmacher 1978). The speed at which a reef is formed depends on rates of skeletongenesis and consolidation, while destruction rates depend upon physical, chemical and biological eroding processes (Risk and Sammarco 1982, Hutchings 1986).
In a healthy reef, bioerosion is very important since it increases the complex-ity of substrates, constantly creating free spaces for coral larvae and other organisms, thus maintaining diversity in species and renovating populations (Hutchings 1986); however in some zones such as the reef crest of Isla Grande where coral growth is limited, bioerosion can eventually kill the reef.
The purpose of this paper is first to describe the bioeroding cryptic fauna associated to skeletons of dead Acropora palmata, responsible for the bioeroding action. Cryptic fauna means organisms living in small pores, holes or cavities within the rock or inside organisms that cover them (Hutchings 1978, Peyrot-Clausade 1981, Scott 1987). Second, to establish whether differences exist between the cryptobiota inhabiting colony branches in life position and broken fallen fragments.
Materials and Methods
Area: Rosario islands' (1008'-10015' N, 75040'- 75048' W) reef complex is formed by approximately 28 coralline islands, 45 kilometers southwest of Cartagena (Colombia); most of the islands are fringed by reefs and seagrass meadows, which are protected within the Rosario Coral Natural National Park (PNNCR) (Sarmiento et al. 1989). Isla Grande is the largest of the islands; parallel to its northern margin, and separated by a small reef channel, lies a large fringing reef reaching 950 m in length (Penereiro 1988). Initial studies in this zone describe a healthy coralline community (Pfaff 1969, Werding and Sánchez 1979). However, from 1982 onwards, a drastic reduction of coral cover, mainly of Acropora palmata, was detected, reaching a values as less as 0.2% of cover of living tissue in 1988 (Garzón-Ferreira and Kielman 1993). The origin and evolution of the degradating process are not yet well understood.
Methodology: coral fragments covered by filamentous algae were collected in April 1991. Five branches in live position and five fallen fragments, approximate volume of 1dm3 each, of A palmata were evaluated (Brock and Smith 1983, Grajal and Laughling 1984). Each fragment was covered with a tightly sealed cotton bag, then broken into small pieces with hammer and chisel over a thick blanket, preventing any loss of material. Macroscopic organisms were narcotized for six hours using magnesium sulfate, then fixed in 10% formaldehyde neutralized with borax and finally preserved in alcohol or formaldehyde, depending from the organims. The volume of each fragment was determined by water displacement (Young 1986).
The sediments and material contained in the bags, as well as the remains obtained after macroscopic sorting with hammer and chisel were meshed with a 100 micron mesh. The resulting material was observed with a stereoscope and the organisms found were preserved in 70% alcohol.
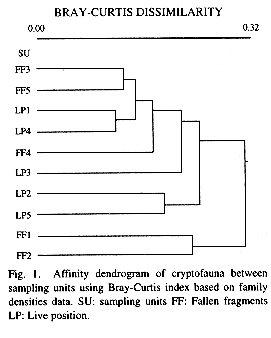
Mathematical analysis of macrocryptobiota was performed on a family level, the lowest taxonomic unit common to all organisms found. Bray-Curtis dissimilarity between fragments was calculated with density data (number of individuals per dm3).
As an approximation to the total biomass (epibiont and cryptobiont), six pieces of coral were fragmented, then dried and calcinated until constant weight was reached. Results were expressed as percent of a coral's dry weight.
Results
In a transversal cut of a branch of A. palmata three horizons can be clearly distinguished. The superior conformed by a film of filamentous algae and its associated organisms. Then, the middle part of the branche, that is skeleton, and the inferior horizont, formed by a crust of coralinaceous algae.
Then, dead Acropora palmata's skeletons supports three types of organisms:
Mobile epibenthon: Composed by the fauna obtained from the examined sediments, living in associations over and under coralline surfaces covered by filamentous algae. Most abundant were small crustaceans, micropolychaetes and foraminifers. Gammarids and caprellids were common amphipods both found in all states of development, tanaidacea was very common and frequently found with eggs. Concerning isopoda, asellots were the most abundant in number while most of the species were flabellifers (Dinamenella, Geocerceis and Exocoralla genus). The most commonly found foraminifers belong to the genera Triloculina and Qinqueloculina, (Myliolidae) although individuals of the Cyclogyra planorbis, Articulina mucronata and Sphaeroina bulloides were also frequent. The Nonionella, Rosalina, Bolivina, and Gipsina genera were also well represented. Associated molluscs Tricolia affinis, Anachis pulchela, A. obesa, Volvarina sp., Hyalina pallida, Cyclostremiscus sp. and various species of Caecum were the species commonest found. It´s acute;s important to notice the presence of the gastropod Marissa cornuaretis, a freshwater mollusc frequently found in the fragments; mainly as empty juvenile shells that are possibly brought to the sea attached to the roots of "taruyas", Eichornia crassipes. This "taruyas" come from the Dique channel, which lies 45 kilometers apart from the Rosario islands, and that connects the Magdalena river with the Cartagena bay.
Boring microcryptobiota: Just underneath the growth of crusty coralinaceous algae, vermetids, and the base of filamentous algae, which penetrated the skeletal matrix, a thin band of perforation was seen. In these horizons evidence of anaerobic organic matter decomposition was found.
Macrocryptobiota: The central skeleton showed varying thickness depending on age and site where the colony was extracted. This skeleton seemed perforated by tubes and holes in an random manner. Cryptobiota, localized in the body of the coralline skeleton were the organisms quantitatively evaluated, measuring 5 millimeters or more. Polychaets were the most abundant followed by sipunculids, mollusks and crustaceans (Table 1), when 7574 individuals were counted and identified in 12.35 dm3 of sample.
Polychaeta reaching a density of 328 individuals per dm3 of coralline rock. Most abundant families were Eunicidae (185 I/dm3) and Nereidae (123 I/dm3). The Cirratulidae, Phyllodocidae, Polynoidae, Dorvilleidae, Terebelidae and Serpulidae families showed low densities. Furthermore there were fifteen types of polychaetes with few individuals that were not identifiable. Polychaetes were frequently associated to clionid sponges and foraminifers, a common situation in these habitats (McCloskey 1970, Lipps and Ronan 1974).
The second largest component in number of organisms were the sipunculids, reaching a density of 213 I/dm3 (Table 1). The population was formed mainly by three genera. Aspidosiphon, type 2, and Phascolosoma, present in all fragments. Behavior is similar to that of polychaetes. Big populations were always present in the fragments but with large variations between them. Various sipunculid perforations communicated directly with the galleries of Alpheus schmitti, and not to the exterior. This was a frequent finding.
The vermetid Dendropoma corrodens is the most abundant mollusc (89 I/dm3) found in the fragments extracted from the north reef crest of Isla Grande (Table 1). Bivalves were the next group in abundance. Caves belonging to bivalves showed intensive infestation by Siphonodyction coralliphagum sponge. Other mollusc individuals were poorly represented in the samples. These were Fissurella angusta, and chitons like Acanthochitona sp., Ischnochiton striolatus and Choneplax lata, of scraping habits. Others were the filtrating bivalves Isognomon bicolor, I. radiatus and free living gastropods such as Engina turbinella and Leucozonia ocellata.
We collected a total of 255 macrocrustaceans distributed in seven families and twelve species (Table 1). Eight Thunor simus cavities in the superior part of a 14 x 14 cm rock were found, each with an eigth shape. They measured 1.32cm maximum mean width and 3.42cm in length. Each of the chambers had a pair of adults; females, always with eggs, and using the larger chamber while the males the smaller. These cavities had no galleries or tunnels interconnecting them, and organisms appear to be completely covered by the skeletal crust of corallinaceous algae.
The most common boring species found in the Acropora palmata scaffold were Cliona cf. celata, Cliona spp., Siphonodictyon coralliphagum, Geodia cf. gibberosa and Aka sp. At a glance of dead branches, presence of any organism is nothing but conspicuous, for their openings to the exterior are through tiny pores. The biggest observed perforations belonged to Geodia cf. giberosa reaching a length of 5 centimeters. Those of Siphonodictyon coralliphagum were of intermediate size and Cliona sp. were the smallest.
Anemones of the Lebrunia coralligens and several species of fish were also found. Some sampling showed a Gobiessocidae Gobiesox punctulatus and the blennoid Hypleurochilus springeri; another two samples had an Acanthemblemaria aspera, and A. rivasi respectively, all of them inhabiting cavities built by other organisms (Greenfield and Greenfield 1982)
Representativity evaluation, based on density data of different families, showed that an acceptable sample is that of four sampling units of one dm3 each. A dissimilarity dendrogram between fragments from both coral positions (Fig. 1) done according to quantified family densities, showed no difference between macrocryptobiota inhabiting live position and fallen fragment corals.
The obtained biomass values (Table 2) were similar throughout except the variation coefficient (80% to 88%).
Discussions
The three kinds of described organisms are found in dead coral blocks (Brawley and Adey 1981, Rice and McIntyre 1982, Grajal and Laughling 1984, Scott and Risk 1988) as well as in rocky shores and submerged rocks, and are not restricted to this habitat and to this particular species (McCloskey 1970, Hutchings 1983, Marshall 1983), but are less abundant than those occurring in the Pacific ocean and in other classes of corals (Patton 1974, Reed and Mikkelsen 1987, Coles 1980).
Mobile epibenthon: Organisms found here (microcrustaceans, micromolluscs and foraminifera) are the most common elements of such habitats (Bell and Deblin 1983, Hutchings 1983, Klumpp et al. 1988). This order of abundance changes between reefs and zones, and depends upon biological cycles of the species (Klumpp et al. 1988)
These abundant kinds of organisms plays a very important roll acting as links between filamentous algae, very abundant in this kind of habitats (Navas et al. in press) and important as primary productors, and reef secondary consumers as fishes, crustaceans larvae and criptobiontes, etc (Brock and Smith 1983).
Boring microcryptobiota: Marshall (1983) and Hutchings (1986) showed that this band was formed by microperforations made by bacteria, fungus, cyanophitic algae, clorophytes and rodophytes and sponges. The predominant species of algae belongs to the genera Hyella, Mastigocoleus, Plectonema, Ostreobium, Phaeophila and Porphyra in caribbean (May et al. 1982).
These organisms are microperforators that allow the entrance of others due to their capacity of chemical dissolution of coral skeletons and are solid bases for microbenthic nourishment (Hutchings 1986 and May et al. 1982).
Macrocryptobiota: First of all we must consider that fauna diversity and abundance is seriously affected by:
1. Hammer and chisel method destroys an important amount of organisms.
2. Some very abundant individuals such as polychaetes were counted on a family level.
3. Organisms with few individuals impossible to identify were not considered for the quantitative analysis.
Classification analysis showed that there was no differences between the two coral positions (live and fragmented) this could happen because there is no differential colonization due to localization, or because the collected fallen fragments were not fixed to substrata so probably they had recently fallen from their branches.
The results of the homogeneity of the fragments and the reduced volume of coralline rock needed to evaluate the community were valid only on a family level. Variations of biomass, density and number of identified organisms on a more specific ground were too ample. This difference between the fragments was due to multiple factors such as:
1. The characteristic position of each fragment plays an important role in the determination of the type of cryptobiont larvae that colonize (Obrebski 1979, Hutchings 1981).
2. The monopolization of various sectors of the substratum by species of closed recruitment and asexual reproduction (Hutchings 1978).
3. Morphologic characteristics of each branch, thickness in particular, seem inversely related to the density of perforating organisms.
4. The type of epibiota, which favors, for filamentous algae, or reduced, for incrusting or carnivorous organisms, the settlement of the eroding larvae (Klumpp et al. 1988).
5. Territorial behavior of fish like Microspathodon chrysurus and Stegastes dorsopunicans. They are abundant in the crest, and protect sections of the coral where filamentous algae grow. This at last favor cryptofaunal colonization, particularly by sponge and sipunculids due to a probable increase in larval settlement, reduction in herbivory and an increase in food supply (Risk and Sammarco 1982, Wins-ton and Jackson 1984).
6. Pioneering colonization during dry or wet season, which determines the ulterior composition of the community (Coles 1980, Hutchings 1981).
The biomass values showed intrinsic differences between each piece of coral, random location of individuals and their occasional presence give variable results.
Acropora palmata mortality in Isla Grande occurred in mass between 1982 and 1983, probably in association with high temperatures (Solano et al.1992). Until 1992 there has not been a significant coral recolonization. What we have observed, though, during this survey, is the result of bioerosion and colonization processes after a period of approximately 8 years. In this succession stage and even though sponges population was not quantified, they seemed mostly responsible for bioerosion of Acropora palmata skeletons in Isla Grande. These organisms are the most important live or dead coral macroborers (Highsmith 1980, Moran and Reaka 1988). The present organisms community, specially polichaetes, according to Davies and Hutchings (1983), are important features in reefs under active degradation processes.
In the other hand some of the secondary colonizers such as vermetids Dendropoma. corrodens, coralline algae and some sponges "fix" coral branches (Wilkinson 1983, Marshall 1983) and make the bioerosion process slower.
Finally, if antropogenic stressers such as pollution, tourism, high temperatures, freshwater inputs and sedimentation keep on acting over Rosario Islands' reefs and avoiding recolonization in affected areas, reef structures such as that found in the north crest of Isla Grande are sentenced to disappear.
Acknowledgements
Special thanks to Clara Osorio for field activities support, Camilo Garcia for his help with statistic software, Rafael Lemaitre and J. M. Diaz for their help with identification and confirmation of species.
Resumen
La criptobiota de diez fragmentos coralinos muertos de Acropora palmata, de 10 dm3 cada uno, cubiertos de algas filamentosas, se colectó en abril de 1991en la cresta arrecifal de Isla Grande (Caribe colombiano). Se halló tres grupos: 1) móviles epibentónicos asociados a las algas filamentosas y conformados principalmente por microcrustáceos; 2) microcriptobiontes perforantes, ubicados en la capa intermedia entre los organismos epilíticos y el esqueleto del coral y 3) macrocriptobiontes que perforan todo el cuerpo del esqueleto coralino (principalmente poliquetos, sipuncúlidos, moluscos y crustáceos). No se encontraron diferencias en la composición de los macrocriptobiontes que habitan los corales en posición de vida y los fragmentos caidos sobre el fondo. Se presentó una amplia variación en biomasa total, tipo y densidad de macrocriptobiontes, posiblemente asociada a factores estocásticos tales como la ubicación y grosor de las ramas, y variaciones del reclutamiento a pequeña escala.
References
Bell S.S. & D.J. Devlin. 1983. Short-term macrofaunal recolonization of sediment and epibenthic habitats in Tampa Bay, Florida. Bull. Mar. Sci. 33: 102-108. [ Links ]
Brawley S.H. & W.H. Adey. 1981. The effect of micrograzers on algal community structure in a coral reef microcosm. Mar. Biol. 61: 167-177. [ Links ]
Brock R.E. & S.V. Smith. 1983. Response of coral reef cryptofaunal communities to food and space. Coral Reefs 1: 179-183. [ Links ]
Coles S.L. 1980. Species diversity of decapods associated with living and dead reef coral Pocillopora meandrina. Mar. Ecol. Prog. Ser. 2: 281-291. [ Links ]
Davies P.J. & P.A. Hutchings. 1983. Initial colonization, erosion and accretion on coral substrate: Experimental results, Lizard Island, Great Barrier Reef. Coral Reefs 2: 27-35. [ Links ]
Garzón-Ferreira J. & M. Kielman. 1993. Extensive mortality of corals in the Colombian Caribbean during the last two decades. Proccedings of the Colloquium on Global Aspects of Coral Reefs: Health, hazards and history. Mar. Atm. Sci. A15-A21. [ Links ]
Grajal A & R. Laughlin. 1984. Decapod crustaceans inhabiting live and dead colonies of three species of Acropora in the Roques Archipiélago, Venezuela. Bijdragen tot de Dierkunde 54: 220-230. [ Links ]
Greenfield D.W. & T.A. Greenfield. 1982. Habitat and resource partitioning betwen two species of Acanthemblemaria (Pisces: Chaenopsidae), with comments on the chaos hypothesis. Smith. Contr. Mar. Sci. 12: 499-507. [ Links ]
Highsmith R.C. 1980. Geographic patterns of coral bioerosion: a productivity hypothesis. J. exp. mar. Biol. Ecol. 46: 177-196. [ Links ]
Hutchings P. 1978. Non-colonial cryptofauna, p. 251-261. In D.R. Stoddart & R.E. Johannes (eds.). Coral reefs research methods, UNESCO, Monograph 19, Paris. [ Links ]
Hutchings P. 1981. Polychaete recruitment onto dead coral substrates at Lizard Island. Great Barrier reef, Australia. Bull. Mar. Sci. 31: 410-423. [ Links ]
Hutchings P. 1983. Cryptofaunal communities of coral reefs, p. 200-208. In D.J. Barnes (ed.). Perspectives on coral reefs. The Australian Institute of Marine Science, Sydney. [ Links ]
Hutchings P. 1986. Biological destruction of coral reefs. Coral Reefs 4: 239-252. [ Links ]
Klumpp D., A. McKinnon & C. Mundy. 1988. Motile cryptofauna of a coral reef, abundance, distribution and trophic potential. Mar. Ecol. Prog. Ser. 45: 95-108. [ Links ]
Lipps J.& T. Ronan. 1974. Predation on foraminifera by the polychaete worm, Diopatra. J. Foraminifera Res. 4: 139-143. [ Links ]
Marshall J. 1983. The fate of borings relations between bioerosion, internal sediment infill and submarine cementation. Proc. Great Barrier Reef Conference, Townsville, Australia, 129-134. [ Links ]
May J.A., I.G. Macintyre & R.D. Perkins. 1982. Distribution of microborers within planted substrates along a Barrier Reef Transect, Carrie Bow Cay, Belize. Smith. Contr. Mar. Sci. 12: 93-107. [ Links ]
McCloskey L.R. 1970. The dynamics of the community associated with a marine scleractinian coral. Int. Revue. ges. Hydrobiol. 55: 13-81. [ Links ]
Moran D.P. & M.L. Reaka. 1988. Bioerosion and availability of shelter for benthic reef organisms. Mar. Ecol. Prog. Ser. 44: 249-263. [ Links ]
Navas G.R., S.K. Moreno-Forero, O.D. Solano & G. Díaz-Pulido. 1998. Ensamblajes arrecifales epiliticos del coral Acropora palmata muerto, isla Grande, Islas del Rosario, Caribe Colombiano. Carib. J. Sci. (in press). [ Links ]
Obrebski S. 1979. Larval colonizing strategies in marine benthic invertebrates. Mar. Ecol. Prog. Ser. 1: 293-300. [ Links ]
Patton W.K. 1974. Community sructure among the animals inhabiting the coral Pocillopora damicornis at Heron island, Australia, p. 219-243. In W. Vernbery (ed) Simbiosis in the sea. University of South Carolina, Columbia. [ Links ]
Penereiro J.L. 1988. Cartografía de mosaicos para fondos submarinos, 409-416. VI Seminario de Ciencias y Tecnologías del Mar, CCO, Bogotá, Colombia. [ Links ]
Peyrot-Clausade M. 1981. Motile cryptofauna of Tulear Great Reef outer slope: Brachyura and Anomura distribution. Proc. Fourth Int. Coral Reef Symposium, Manila 2: 745-754. [ Links ]
Pfaff R. 1969. Las Scleractinia y Milleporina de las Islas del Rosario. Mitt. Int. Colombo-Alemán Invest. Cient. 3: 17-24. [ Links ]
Reed J & P. Mikkelsen. 1987. The molluscan community associated with the scleractinian coral Oculina varicosa. Bull. Mar. Sci. 40: 99-131. [ Links ]
Rice M. & I. Macintyre. 1982. Distribution of sipuncula in the coral reef community Carrie Bow Cay, Belize. Smith. Cont. Mar. Sci. 12: 311-320. [ Links ]
Risk M.J. & P.W. Sammarco. 1982. Bioerosion of corals and the influence of damselfish territory. Oecologia 52: 376-380. [ Links ]
Sarmiento E, F. Flechas & G. Alvis. 1989. Evaluación cuantitativa del estado actual de las especies coralinas del Parque Nacional Natural Corales del Rosario (PPNCR), Cartagena (Colombia). Tesis B. Sc. Biologia Marina, Universidad Jorge Tadeo Lozano, Bogotá, Colombia. [ Links ]
Scott P.J. 1987. Associations between corals and macroinfaunal invertebrates in Jamaica, with a list of Caribbean and Atlantic Coral associates. Bull. Mar. Sci. 40: 271-286. [ Links ]
Scott P.J.B. & M.J. Risk. 1988. The effect of Lithophaga (Bivalvia: Mytilidae) bore holes on the strength of the coral Porites lobata. Coral Reefs 7: 145-151. [ Links ]
Schuhmacher H. 1978. Arrecifes coralinos: Su extensión, mundo animal y ecología. Omega, Barcelona, 288 p. [ Links ]
Solano O.D., G.R. Navas & S.K. Moreno-Forero. 1992. Blanqueamiento coralino de 1990 en el Parque Nacional Natural Corales del Rosario (Caribe Colombiano). An. Inst. Invest. Mar. Punta Betín 22 : 97-111. [ Links ]
Werding B & H. Sánchez. 1979. Situación general y estructuras arrecifales: Informe Faunístico y Florístico de las islas del Rosario en la costa norte de Colombia. An. Inst. Inv. Mar. Punta Betín 11: 7-20. [ Links ]
Wilkinson C.R. 1983. Role of sponges in coral reef structural processes, 263-274 In: Barnes DJ (Ed) Perspectives on coral reefs. The Australian Institute of Marine Science, Sydney. [ Links ]
Winston J.E. & J.B.C. Jackson. 1984. Ecology of cryptic coral reef communities. IV. Community development and life histories of encrusting cheilostome bryozoa. J. Exp. Mar. Biol. Ecol. 76: 1-21. [ Links ]
Young P. 1986. Análise qualitativa e quantitativa da fauna associada a corais hermatípicos (Coelenterata, Scleractinia) nos recifes de Joâo pessoa, PB. Rev. Brasil. Biol. 46: 99-126. [ Links ]
1 INVEMAR. A.A. 1016 Santa Marta, Colombia. e-mail: colref@invemar.edu.co

