











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.46 no.1 San José mar. 1998
Abstract
The effect of nutrient enrichment (N and P) on the phytoplankton community of La Mariposa reservoir (Venezuela) was studied. Polyethylene bags (microcosms) were filled with 10 l of filtered epilimnetic water of the reservoir, and enriched with different nutrient proportions, simulating an eutrophication process. Chlorophyll 'a', total nitrogen, total phosphorus, and phytoplankton abundance were determined. There was an increase in phytoplankton abundance because of nutrient enrichment, specially when both nutrients were present. After the experiments, Cryptophyta, Euglenophyta, and Pyrrophyta dissappeared from the microcosms, whereas Chlorophyta and Cyanobacteria increased. Numeric proportions of diatoms decreased at higher nitrogen concentration. Although the results obtained with microcosm experiments could be limited in ecological significance, they permit an useful approach to the natural situation.
Key words
Microcosm, nutrient enrichment, eutrophication, La Mariposa Reservoir, Venezuela.
Los cambios estacionales en la disponibilidad de nutrientes, especialmente nitrógeno y fósforo, pueden causar cambios en la estructura comunitaria del fitoplancton (Tundisi y Henry 1986, Vanni y Temte 1990).
El enriquecimiento con nutrientes se ha convertido en un problema a partir de la última mitad del siglo XX, ya que ha generado la eutroficación de numerosos cuerpos de agua (Harper 1992).
En Venezuela son escasos los trabajos relacionados con el efecto del enriquecimiento de nutrientes sobre la comunidad del fitoplancton (González et al. 1991, González 1996), al igual que en el trópico suramericano (Henry y Tundisi 1982, Tundisi y Henry 1986).
En vista de lo anterior, se planteó el objetivo de estudiar el efecto del enriquecimiento con nitrógeno y fósforo sobre la comunidad del fitoplancton del embalse La Mariposa, y evaluar las respuestas iniciales de esta comunidad ante un proceso de eutroficación. Este embalse soporta una fertilización excesiva debido al crecimiento de asentamientos humanos en su cuenca, lo cual puede afectar el uso del que es objeto en la actualidad (como suministro de agua potable a la población).
Materiales y Métodos
El embalse La Mariposa está ubicado a 8 km de la ciudad de Caracas (10o24'41"N; 66o33'53" W) a 980 m.s.n.m. Este embalse represa al río El Valle y la quebrada Los Indios con la finalidad de abastecer de agua potable a la ciudad de Caracas; sin embargo la mayor proporción de agua que recibe proviene del bombeo desde otros embalses, como los de Camatagua y Lagartijo.
El volumen de agua embalsada es de 4 000 000 m3, cubre un área de 600 000 m2 y las profundidades máxima y media son 20 m y 6.7 m, respectivamente (Vivas 1988). En la actualidad, los afluentes del embalse están contaminados con materia orgánica, provenientes de las actividades de caseríos, cochineras y viveros que operan en la cuenca de drenaje.
En una estación cercana al dique, se tomaron muestras de 10 l de agua epilimnética. Para evitar la interferencia en el experimento, los organismos del zooplancton se excluyeron mediante el filtrado de las muestras a través de una red con abertura de malla de 80 ![]() m antes de ser colocadas en cada una de las bolsas plásticas transparentes o microcosmos (30 cm de diámetro y 70 cm de longitud). Las bolsas fueron lavadas previamente con HCl 10 % y enjuagadas con agua corriente y agua del embalse.
m antes de ser colocadas en cada una de las bolsas plásticas transparentes o microcosmos (30 cm de diámetro y 70 cm de longitud). Las bolsas fueron lavadas previamente con HCl 10 % y enjuagadas con agua corriente y agua del embalse.
El agua filtrada se sometió a diversos tratamientos con adición de nutrientes (cloruro de amonio y dihidrógeno fosfato de potasio), sugeridos por Elser y Goldman (1991) (Cuadro 1). Para efectos de comparación con los resultados registrados para ambientes tropicales, las concentraciones finales se expresaron en ![]() g/l. Se realizaron 9 tratamientos, cada uno con su respectivo duplicado. Las muestras permanecieron suspendidas en la superficie del embalse entre el 23 y el 29 de febrero de 1996.
g/l. Se realizaron 9 tratamientos, cada uno con su respectivo duplicado. Las muestras permanecieron suspendidas en la superficie del embalse entre el 23 y el 29 de febrero de 1996.
| Fósforo | |||
| (KH2PO4 |
| | |
| 0 | | | |
| 310 | | | |
| 620 | | | |
Al inicio del experimento se tomaron muestras de agua de la misma zona para estimar las concentraciones iniciales de fósforo total y nitrógeno total (Valderrama 1981), de clorofila 'a' (Nusch y Palme 1975) y la abundancia numérica del fitoplancton (Wetzel y Likens 1991). Además, se estimaron la transparencia del agua (disco de Secchi de 20 cm de diámetro) y la conductividad eléctrica (S/cm).
Al final del experimento se estimaron, para cada tratamiento, la concentración de clorofila 'a', las concentraciones de N total y P total y la abundancia numérica del fitoplancton.
Los valores de clorofila 'a' para cada tratamiento se contrastaron con un ANOVA de dos vías (p<.05), luego de verificar las condiciones para su aplicación. Para comparar si hubo diferencias significativas entre el orden por rangos de las abundancias de los diferentes grupos del fitoplancton al inicio y al final de cada experimento, se aplicó una correlación no paramétrica de Spearman (rS, p<.05).
Resultados
Las condiciones del embalse, al inicio de los experimentos, fueron las siguientes:
El color del agua era verde oliva, y la transparencia de 0.3 m. La conductividad eléctrica fue 225![]() S/cm, lo cual permite clasificarlo como de baja salinidad, según Serruya y Pollingher (1983).
S/cm, lo cual permite clasificarlo como de baja salinidad, según Serruya y Pollingher (1983).
La concentración de fósforo total fue de 28.5 ![]() g/l, mientras que la de nitrógeno total fue de 2 523.0
g/l, mientras que la de nitrógeno total fue de 2 523.0 ![]() g/l, con una relación N: P de 88.5: 1, lo cual sugiere que el nutriente limitante para el crecimiento del fitoplancton es el fósforo. Con base en la concentración de fósforo total, el embalse La Mariposa puede clasificarse, según el índice de Salas y Martinó (1990), como mesotrófico.
g/l, con una relación N: P de 88.5: 1, lo cual sugiere que el nutriente limitante para el crecimiento del fitoplancton es el fósforo. Con base en la concentración de fósforo total, el embalse La Mariposa puede clasificarse, según el índice de Salas y Martinó (1990), como mesotrófico.
La concentración inicial de clorofila 'a' fue de 32.0 ![]() g/l.
g/l.
En las muestras analizadas no se encontraron organismos del zooplancton, indicando que la exclusión con la red de abertura de malla de 80 ![]() m fue efectiva.
m fue efectiva.
Al final del experimento, las concentraciones de N y P en el tratamiento sin enriquecer fueron las mismas que aquéllas encontradas en el ambiente natural ("ambiente") (Fig. 1).
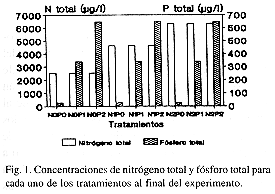
Con excepción de los tratamientos N0P1, N0P2 y N1P2, donde la proporción N: P fue menor que 9, en todos los tratamientos hubo limitación por el P. En el ambiente y en los tratamientos sin enriquecer con P (P0), la relación N: P fue mayor que 50: 1, lo que podría indicar una severa limitación por el fósforo (Fig. 2).
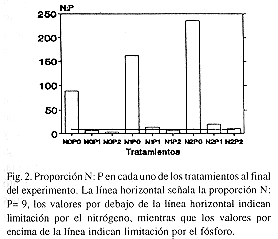
En los tratamientos en los que ambos nutrientes estuvieron ausentes (N0P0) o sólo uno de ellos estuvo ausente (N0P1, N0P2, N1P0, N2P0), las concentraciones de clorofila 'a' resultaron menores que en los casos en los que ambos nutrientes estuvieron presentes (Fig. 3).
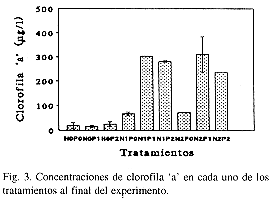
La respuesta al enriquecimiento con nitrógeno solamente (N1P0 y N2P0) fue más pronunciada que la obtenida con fósforo solamente (N0P1 Y N0P2), resultando en estos últimos casos las concentraciones de clorofila 'a' muy parecidas al tratamiento sin enriquecer con nutrientes (N0P0). Es posible que, a pesar de las altas concentraciones de N en el embalse, este elemento no se encontraba en forma disponible para los organismos del fitoplancton durante el inicio de los experimentos.
Un ANOVA de dos vías demostró que hubo diferencias significativas entre los tratamientos enriquecidos con ambos nutrientes y aquéllos en los que uno de los nutrientes o ambos estaban ausentes (Cuadro 2). Sin embargo, no hubo diferencias significativas entre la aplicación de una dosis "doble" de nutrientes (N2, P2) y la de una dosis "sencilla" (N1, P1). La prueba "a posteriori" (diferencia mínima significativa), demostró que los microcosmos enriquecidos resultaron ser significativamente diferentes de los no enriquecidos.
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | | | |
| | |
En cuanto a las proporciones del fitoplancton en el ambiente, Bacillariophyta fue el grupo dominante, representando más del 50 % de los organismos presentes. Aulacoseira pseudogranulata fue la especie dominante al inicio de los experimentos. Otras especies de diatomeas presentes fueron Cyclotella meneghiniana, Fragilaria sp. y Navicula sp. El grupo de las Cryptophyta, representadas por Cryptomonas erosa, fue el segundo en dominancia en el ambiente, seguido por las Cyanobacteria (Anabaenopsis raciborskii, Chroococcus sp., Dactylococcopsis acicularis, Lyngbya limnetica, Microcystis aeruginosa, Oscillatoria sp., Spirulina sp. y Synechococcus sp.). Las Chlorophyta (Actinastrum hantzschii, Dictyosphaerium pulchellum, Oocystis sp., Pediastrum simplex, Scenedesmus ecornis, Sphaerocystis schroeteri, Staurastrum sp. y Schroederia sp.) representaron el 3.8 % del total, mientras que las Euglenophyta (Trachelomonas sp.) y las Pyrrophyta (Peridinium sp.) no alcanzaron el 2 % del total (Fig. 4).
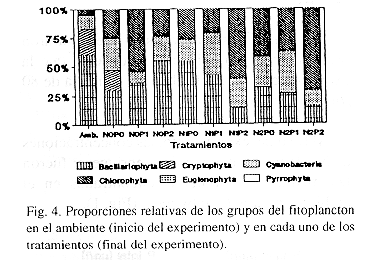
Las condiciones de encierro en los microcosmos, afectaron a los diferentes grupos del fitoplancton. Al término del período de experimentación, al comparar el tratamiento sin enriquecer (N0P0) con la condición inicial ("ambiente") (Fig. 4), se pudo notar que los géneros representantes de Pyrrophyta y de Euglenophyta desaparecieron del sistema experimental, la proporción de Bacillariophyta disminuyó mientras que aumentaron las proporciones de Chlorophyta y de Cyanobacteria. La proporción relativa de Cryptophyta se mantuvo en los mismos niveles al finalizar el experimento. Los valores de rS obtenidos indicaron diferencias significativas en las proporciones numéricas de los distintos grupos del fitoplancton al comparar el ambiente con los distintos tratamientos experimentales. El orden de importancia relativa de los grupos del fitoplancton del tratamiento sin enriquecer (N0P0) estuvo correlacionado significativamente con el de los tratamientos enriquecidos N1P0 y N1P1 (rS= 0.941 y rS= 0.955, respectivamente).
En general, se observó que la fertilización con nutrientes trajo como resultado la desaparición de los grupos Cryptophyta, Euglenophyta y Pyrrophyta de los sistemas experimentales (Fig. 4). La proporción relativa de las Chlorophyta, comparada con la del ambiente natural, aumentó en todos los microcosmos y los géneros más abundantes fueron Schroederia, Sphaerocystis y Scenedesmus.
Las proporciones relativas de las Cyanobacteria aumentaron en los microcosmos con respecto al "ambiente", excepto en los tratamientos N0P1 y N2P2, donde permanecieron casi iguales que al inicio del experimento. Microcystis fue el género dominante de este grupo, seguido por Anabaenopsis.
Las Bacillariophyta fueron dominantes en los tratamientos sin enriquecer con nitrógeno (N0) y con dosis sencilla de nitrógeno (N1), excepto en el tratamiento N1P2. En los tratamientos con dosis doble de nitrógeno (N2), este grupo presentó las menores proporciones relativas. El género dominante de este grupo fue Aulacoseira.
Discusión
Los ambientes experimentales empleados en este trabajo pueden considerarse como instrumentos para estudiar las propiedades de los ecosistemas en condiciones bien definidas, de manera que las observaciones puedan ser obtenidas con mejor calidad estadística que la posible en el campo (Ringelberg y Kersting 1978). Este tipo de "microecosistemas" pueden tener un valor realístico suficiente, especialmente si son de vida corta (Ringelberg y Kersting 1978, Matveev 1991). Havens y De Costa (1986) también comentan que el encierro "per se" altera el ambiente dentro de las bolsas cuando se compara con el lago, aunque los datos químicos de estos ambientes experimentales no lo demuestren. Esto se debe a que los ambientes dentro de las bolsas son estáticos, ya que impiden la entrada de nutrientes. Esto explicaría la falta de una correlación entre el orden por rangos de los grupos del fitoplancton al inicio del experimento y el tratamiento N0P0.
Según Edmondson (1957), el enriquecimiento con nutrientes provoca un rápido aumento en las concentraciones de clorofila 'a', lo cual coincide con lo encontrado en esta experiencia. Estos resultados son similares a los obtenidos por otros investigadores al trabajar con encierros de comunidades del fitoplancton en bolsas plásticas o en limnocorrales, entre los que se pueden citar De Costa et al. (1983), Bergquist y Carpenter (1986), Pollingher et al. (1988), Elser y Goldman (1991), Yasunoet al. (1993), entre muchos otros.
Henry y Tundisi (1982) y Tundisi y Henry (1986), estudiaron el efecto del enriquecimiento con nutrientes en limnocorrales en sistemas tropicales (embalse de Broa y lago Dom Helvécio, Brasil). Al igual que en otros estudios, la adición de nutrientes aumentó la biomasa del fitoplancton, medida como concentración de clorofila 'a'. Estos investigadores encontraron que el fósforo tuvo un rol más significativo en el aumento de la clorofila 'a' que el nitrógeno y otros micronutrientes. Estos mismos resultados fueron obtenidos por González (1996) en microcosmos en el embalse El Andino, Venezuela, quien encontró una correlación positiva significativa entre el fósforo total y las concentraciones de clorofila 'a', pero no encontró correlación entre la biomasa del fitoplancton y el nitrógeno total.
En el caso de los microcosmos del embalse La Mariposa, a diferencia de los ejemplos anteriores, se encontró que la biomasa del fitoplancton respondió en mayor escala a los tratamientos con nitrógeno que a los tratamientos con fósforo. Estos resultados coinciden con los obtenidos por Cowell y Dawes (1991), quienes encontraron que el nitrógeno (en forma de amonio) produjo aumentos significativos de la clorofila 'a' en todos sus experimentos de bioensayos en tres lagos de diferentes estados tróficos en Florida, Estados Unidos. En el presente estudio, en los tratamientos en los que ambos nutrientes estaban presentes, el aumento de la concentración de clorofila 'a' fue más pronunciado que en los casos en que el N o el P estaban ausentes. Esto quedó reflejado en la prueba estadística aplicada (ANOVA), al resultar estadísticamente significativa la interacción entre los nutrientes (N x P, Cuadro 2).
En el ambiente, la alta proporción N: P ayudaría a explicar la dominancia de las diatomeas en esta época del año, ya que varias especies de este grupo tienen bajos coeficientes de saturación media para el fósforo, lo cual las hace superiores competitivamente sobre especies de otros grupos cuando este elemento es limitante (Margalef 1983). La alteración de estas relaciones por el encierro de la comunidad en bolsas de polietileno, pudiera explicar las variaciones de las proporciones de las Bacillariophyta con respecto a los otros grupos del fitoplancton. La conducta de unas especies en una trama de variables interactuantes en la naturaleza es completamente diferente a su comportamiento bajo condiciones controladas (Bloesch et al. 1988).
Sommer (1983, 1988) encontró que Scenedesmus (Chlorophyta), es una especie típica en las etapas tempranas de una sucesión luego del enriquecimiento con nutrientes. Scenedesmus se encontró comúnmente al final de los experimentos con los microcosmos en el embalse La Mariposa. Al igual que los resultados obtenidos por Sommer (1988), los flagelados como Cryptomonas no fueron especies exitosas y desaparecieron de los sistemas experimentales. Lo mismo sucedió con Peridinium sp. y con Trachelomonas sp.
Blomqvist et al. (1994) señalan que las Cyanobacteria no fijadoras de N proliferan cuando la fuente de nitrógeno está en forma de cloruro de amonio, tal como la empleada en este trabajo. Esto coincidió con el aumento de la proporción de Cyanobacteria en todos los microcosmos con respecto a su proporción en las aguas del embalse La Mariposa, excepto en los tratamientos N0P1 y N2P2.
Generalmente, se sugiere que las Cyanobacteria son mejores competidoras por el nitrógeno, pero pobres competidoras por el fósforo cuando se les compara con otras especies del fitoplancton (Smith 1983, Drenner et al. 1990). Las Cyanobacteria dominarían, por lo tanto, en situaciones en las que la proporción N: P fuera menor que 9. Sin embargo, la proporción N: P por sí sola no es suficiente para explicar la ausencia o la presencia de las diversas especies en los lagos (Smith 1983). Sommer (1990), apunta que las tasas de suministro de N y de P, más que la concentración de estos elementos, son las limitantes del crecimiento de las algas. Esto parece quedar en evidencia en los tratamientos N0P1 y N1P2, con proporciones N: P similares (7.5: 1 y 7.1: 1, respectivamente), pero con concentraciones de clorofila 'a' diferentes. En ambos casos, el nitrógeno parece ser el limitante, pero en N1P2, el exceso de nutrientes generó una diferencia casi veinte veces mayor.
Los resultados obtenidos en los microcosmos pudieran indicar que las respuestas iniciales de la comunidad del fitoplancton del embalse La Mariposa ante un proceso de eutroficación, iniciado en la época del año en la cual se realizó el estudio, serían: 1) el aumento de la biomasa (como concentración de clorofila 'a'); 2) la desaparición de los grupos Pyrrophyta, Cryptophyta y Euglenophyta; 3) el aumento de la dominancia de las Chlorophyta, en primera instancia, y de las Cyanobacteria; y 4) la disminución de la dominancia de las diatomeas, especialmente a altas concentraciones de nitrógeno.
Agradecimientos
A Paula Spiniello, Jorge Gutic, Nora Malaver, Carlos Peñaherrera y los integrantes del curso de Laboratorio de Ecología II (1995-96), por el apoyo en el trabajo de campo y en el laboratorio. A Carlos López y Aída González de Infante, por la revisión y las sugerencias para mejorar el manuscrito.
Resumen
Para evaluar el efecto del enriquecimiento con nutrientes sobre la comunidad del fitoplancton del embalse La Mariposa (Venezuela), se establecieron microcosmos "in situ" durante 6 días. Los microcosmos consistieron en bolsas de polietileno con 10 l de agua epilimnética filtrada del embalse, sometidos a diferentes combinaciones de enriquecimiento con fósforo y nitrógeno, para así simular un proceso de eutroficación. Se determinaron las concentraciones de clorofila 'a', la concentración de nitrógeno total y fósforo total y la proporción numérica de los diferentes grupos del fitoplancton. Como consecuencia del enriquecimiento de nutrientes, se produjo un aumento en la biomasa del fitoplancton, siendo las respuestas más pronunciadas en los tratamientos donde ambos nutrientes estaban presentes. Los grupos Cryptophyta, Euglenophyta y Pyrrophyta desaparecieron de los microcosmos fertilizados al finalizar el período de estudio, mientras que las Chlorophyta y las Cyanobacteria aumentaron sus proporciones relativas. Las diatomeas disminuyeron su proporción relativa a altas concentraciones de nitrógeno. Aunque la técnica del microcosmos tiene limitaciones en su significado ecológico, permite obtener una valiosa aproximación para entender lo que ocurre en las situaciones naturales.
Referencias
Bergquist, A.M. & S.R. Carpenter. 1986. Limnetic herbivory: Effects on phytoplankton populations and primary production. Ecology 67: 1351-1360. [ Links ]
Bloesch, J., P. Bossard, H. Bührer, H.R. Bürgi & U. Uehlingher. 1988. Can results from limnocorral experiments be transferred to in situ conditions? ( Biomanipulation in limnocorrals VI). Hydrobiologia 159: 297-308. [ Links ]
Blomqvist, D.O., A. Pettersson & F. Hyenstrand. 1994. Ammonium-nitrogen: a key regulatory factor causing dominance of non-nitrogen-fixing Cyanobacteria in aquatic systems. Arch. Hydrobiol. 132: 141-164. [ Links ]
Cowell, B. & C.J. Dawes. 1991. Nutrient enrichment in three Central Florida lakes of different trophic states. Hydrobiologia 220: 217-231. [ Links ]
De Costa, J., A. Janicki, G. Shellito & G. Wilcox. 1983. The effect of phosphorus additions in enclosures on the phytoplankton and zooplankton of an acid lake. Oikos 40: 283-294. [ Links ]
Drenner, R.W., J.D. Smith, J.R. Mummert & H.F. Lancaster. 1990. Responses of an eutrophic pond community to separated and combined effects of N: P supply and planktivorous fish: a mesocosm experiment. Hydrobiologia 208: 161-167. [ Links ]
Edmondson, W.T. 1957. Trophic relations of the zooplankton. Trans. Amer. Micr. Soc. 76: 225-245. [ Links ]
Elser, J.J. & C.R. Goldman. 1991. Zooplankton effects on phytoplankton in lakes of contrasting trophic status. Limnol. Oceanogr. 36: 64-90. [ Links ]
González, E., J. Paolini & A. Infante. 1991. Water chemistry, physical features and primary production of phytoplankton in a tropical blackwater reservoir (Embalse de Guri, Venezuela). Verh. Internat. Verein. Limnol. 24: 1477-1481. [ Links ]
González, E.J. 1996. Interacciones fitoplancton-zooplancton en condiciones naturales y experimentales en un embalse de la cuenca del río Unare (embalse El Andino, Edo. Anzoátegui). Tesis de Doctorado, Universidad Central de Venezuela, Caracas, Venezuela. [ Links ]
Harper, D. 1992. Eutrophication of freshwaters. Principles, problems and restoration. Chapman & Hall, Londres. 327 p. [ Links ]
Havens, K.E. & J. De Costa. 1986. A comparison of phytoplankton responses to nutrient addition in acidic and circumneutral pH lakewater. Hydrobiologia 137: 211-222. [ Links ]
Henry, R. & J.G. Tundisi. 1982. Efeitos de enriquecimento artificial por nitrato de fosfato no crescimento da comunidade fitoplanctônica da Represa de Lobo ("Broa", Brotas-Itirapina, SP). Ciência e Cultura 34: 518-524. [ Links ]
Margalef, R. 1983. Limnología. Omega, Barcelona. 1010 p. [ Links ]
Matveev, V. 1991. Self-maintaining plankton: pelagic Cladocera in small microcosms with lake water. Hydrobiologia 225: 301-307. [ Links ]
Nusch, E.A. & G. Palme. 1975. Biologische Methoden für der Praxis der Gewässeruntersuchung, Bestimmung des Chlorophyll-a und Phaeopigment-gehaltes in Oberflachenwässer. GWF-Wässer/ Abwässer 116: 562-565. [ Links ]
Pollingher, U., T. Berman, B. Kaplan & D. Scharf. 1988. Lake Kinneret phytoplankton: response to N and P enrichments in experimental and in nature. Hydrobiologia 166: 65-75. [ Links ]
Ringelberg, J. & K. Kersting. 1978. Properties of an aquatic micro-ecosystem. I. General introduction to prototypes. Arch. Hydrobiol. 83: 47-68. [ Links ]
Salas, H. & P. Martinó. 1990. Metodologías simplificadas para la evaluación de eutroficación en lagos cálidos tropicales. Programa Refioval CEPIS/HPE/IOPS, Lima. 52 p. [ Links ]
Serruya, C. & U. Pollingher. 1983. Lakes of the warm belt. Cambridge University, Cambridge. 569 p. [ Links ]
Smith, V.H. 1983. Low nitrogen to phosphorus ratios favor dominance by blue-green algae in lake phytoplankton. Science 221: 669-671. [ Links ]
Sommer, U. 1983. Nutrient competition between phytoplankton species in multispecies chemostat experiments. Arch. Hydrobiol. 96: 399-416. [ Links ]
Sommer, U. 1988. Phytoplankton succession in microcosm experiment under simultaneous grazing presence and resource limitation. Limnol. Oceanogr. 33: 1037-1054. [ Links ]
Sommer, U. 1990. Phytoplankton nutrient nutrient competition from laboratory to lake, p. 193-213. In: J.B. Grace and D. Tilman (eds.). Perspectives on plant competition. Academic., San Diego. [ Links ]
Tundisi, J.G. & R. Henry. 1986. Effects of enrichment on the summer surface phytoplanktonic community in a stratified tropical lake ( Lake Dom Helvécio - Parque Florestal do Rio Doce Minas Gerais). Rev. Brasil Biol. 46: 231-237. [ Links ]
Valderrama, J.C. 1981. The simultaneous analysis of total nitrogen and total phosphorus in natural waters. Mar. Chem. 10: 109-122. [ Links ]
Vanni, M.J. & J. Temte. 1990. Seasonal patterns of grazing and nutrient limitation of phytoplankton in a eutrophic lake. Limnol. Oceanogr. 35: 697-709. [ Links ]
Vivas, L. 1988. Algunas características físicas y químicas del embalse La Mariposa (Dtto. Federal). Tesis de Licenciatura, Universidad Central de Venezuela, Caracas, Venezuela. [ Links ]
Wetzel, R. & G.E. Likens. 1991. Limnological analyses. 2nd. edition. Springer-Verlag, Nueva York. 391 p. [ Links ]
Yasuno, M., N. Takamura & T. Hanazato. 1993. Nutrient enrichment experiment using small microcosms, p. 181-193. In: Wetlands and ecotones. Studies on land-water interactions. B. Gopal, A. Hillbricht-Ilkowska & R.G. Wetzel (eds.). National Institute of Ecology and National Scientific Publications, Nueva Delhi. [ Links ]
1 Universidad Central de Venezuela, Instituto de Biología Experimental, Apartado 47106 - Los Chaguaramos, Caracas 1041, Venezuela.

