Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.51 n.1 San José Mar. 2003
Seven reef formations were studied in South Caicos, Turks & Caicos, to determine the species richness, patterns of diversity and patterns of distribution of crustaceans and echinoderms, and to evaluate the relationship between these parameters and the degree of sedimentation of the different sites. The reefs showed a gradient from a high sedimentation level, almost totally covered by algae, to places with no sediment particles deposited over the corals. Sites were classified as with high, low or null sedimentation, and species richness, abundance, diversity, spatial distribution of species and similarity among sites were estimated. No unique pattern was found: for crustaceans as well as for echinoderms, the site with the highest diversity value and high equitability, presumably associated to the environmental heterogeneity of this reef formation, showed null sedimentation and an uniform and random pattern of distribution, crustaceans and echinoderms respectively. The two sites with the lowest diversity for both animal groups, although with different sedimentation levels, showed the lowest equitability value and were the only sites with an aggregated pattern of distribution. The next sites in diversity for crustaceans were those with high sedimentation, probably because most species present inhabit empty conchs, in the sediment, or among seagrasses. For the echinoderms, on the contrary, the intermediate sites in diversity had low sedimentation; the habitat requirements for these species (inside sponges, over the corals or among rocks) may have determined this result. The sites with lowest diversity had high sedimentation levels. In these, crustaceans showed the lowest equitability values and an aggregated spatial distribution, while the community of echinoderms was dominated by one single species. Although only general descriptions can be elucidated with the present results, knowledge about the basic population characteristics and natural history of these reef communities, combined with that of the perturbations related to human activities, provides useful base information for appropriate planning for the restoration and conservation of this ecosystem.
Keywords: coral reefs, echinoderms, crustaceans, diversity, sedimentation, community structure, Caribbean.
Los arrecifes de coral son los ecosistemas marinos taxonómicamente más diversos, los cuales proveen un hábitat complejo para un sinnúmero de especies y donde invertebrados, peces, ortugas, algas y otros organismos forman parte integral de la comunidad arrecifal (Goreau et al. 1979, Richmond 1993, Reaka-Kudla 1997, Vázquez-Domínguez 2000). Los arrecifes coralinos se reconocen actualmente como ecosistemas complejos, con múltiples estados estables, fluctuantes en el tiempo (Jackson 1997, 2001, Wood 2001). Asimismo, ya no se asumen como ambientes "prístinos", sino como ecosistemas que han sido modificados y sobre-explotados tanto histórica como recientemente (Jackson 1997, 2001, Nyström y Folke 2001).
Sin embargo, se reconoce también que numerosos factores son importantes para determinar la distribución y diversidad de especies en los arrecifes, y que la mayoría de las especies presentes exhiben un intervalo de tolerancia relativamente estrecho a las condiciones de su hábitat (Kleypas et al. 1999). Son ecosistemas asociados a niveles particulares de diferentes variables ambientales que afectan el crecimiento y la sobrevivencia de los organismos (e.g. temperatura, luz, salinidad, nutrientes; Huston 1985, Richmond 1993, Kleypas et al. 1999). Por otro lado, las mareas, la acción del oleaje, la sedimentación y la actividad de herbívoros y depredadores, son factores que afectan los patrones de sucesión y colonización y son importantes en la regulación de la diversidad de este ecosistema (Nyström y Folke 2001, Wood 2001). Para algunos corales poco resistentes a la depositación de partículas, la sedimentación en particular puede actuar como una perturbación (episódica o crónica), así como un regulador de las tasas de crecimiento del coral, afectando directa o indirectamente a otros invertebrados y vertebrados que componen la comunidad arrecifal (Huston 1985, Rogers 1990, Gleason 1998, Nyström y Folke 2001).
Las características básicas de una comunidad tales como riqueza, abundancia, diversidad y patrones de distribución de las especies, entre otros, continua siendo información indispensable para su adecuado conocimiento. Aunado a esto, es necesario conocer los factores bióticos y abióticos que determinan la estructura de la comunidad misma (De Vantier et al. 1998, Karlson y Cornell 1999, Ezcurra et al. 2001). La diversidad de especies es un patrón ecológico medible a nivel de la comunidad y donde, en general, una alta diversidad refleja una comunidad ecológicamente compleja (Magurran 1988). Los índices de diversidad se utilizan para determinar la riqueza de especies y la diversidad relativa de una comunidad, frecuentemente asociados con datos ecológicos que permiten describir la estructura y complejidad de la misma (Connell 1978, Magurran 1988). Por otro lado, los índices de diversidad han sido ampliamente utilizados en estudios de monitoreo para evaluar cómo cambia la diversidad por efecto de diferentes factores, naturales o artificiales, que impactan a la comunidad. Entre éstos, el índice de Shannon se ha empleado para el monitoreo de comunidades bénticas (Wu 1982, Pointer y Kennedy 1984, Magurran 1988).
Diversos sistemas arrecifales de la región occidental del Atlántico han sufrido cambios en la estructura de sus comunidades en las últimas décadas, sobre todo en el Caribe, donde muchos están dominados por comunidades de macroalgas (Edmunds y Carpenter 2001). Por otro lado, la historia muestra que los ecosistemas costeros del Caribe estaban severamente degradados mucho tiempo antes de que se iniciaran estudios ecológicos en estos ambientes (Jackson 1997, 001). En esta zona, el archipiélago de las Bahamas presenta una historia geológica particular. La base de las Bahamas es parte del piso oceánico recientemente formado sobre la placa Norteamericana y, durante las últimas glaciaciones, los sedimentos sueltos de la zona se cementaron formando lo que actualmente conocemos como la Plataforma de las Bahamas, la cual es relativamente joven (i.e. no mayor de 5 mil años; Sealey 1994). La producción de sedimento es resultado de la producción litogénica y biogénica de carbonato de calcio (i.e. sedimentos formados por procesos físicos de precipitación química exclusivos de este archipiélago y sedimentos formados a partir de elementos esqueléticos de algas e invertebrados, respectivamente). El movimiento de sedimentos es determinado por corrientes que desplazan grandes volúmenes de agua hacia dentro y fuera del archipiélago, vía mareas semi-diurnas Sullivan 1991, Sealey 1994).
Las formaciones arrecifales de South Caicos, en Turks & Caicos, que forman parte de la Plataforma de las Bahamas, varían de manera importante en cuanto al nivel de sedimentación que presentan, determinado por un gradiente que va desde con un alto grado de sedimentación y que se refleja en la predominancia de sustrato arenoso y de comunidades de algas en algunos arrecifes, con evidente escasez de corales y de otras especies asociadas a los mismos, hasta sin partículas de sedimentos depositados sobre los corales. Este gradiente de sedimentación está asociado a diferencias en la energía del oleaje entre los arrecifes cercanos a islas y aquellos más expuestos (Chiappone et al. 1996). El objetivo del presente estudio fue determinar la riqueza, diversidad, abundancia y distribución espacial de crustáceos y equinodermos en formaciones arrecifales de South Caicos. Asimismo, evaluar la relación entre las características de sedimentación en los arrecifes y dichos parámetros poblacionales.
Materiales y métodos
Área de estudio: las islas Turks y Caicos, colonia británica, comprenden la extensión suroriental de la Plataforma de las Bahamas (21-22ºN y 71-72ºW). Archipiélago de piedra caliza que representa la superficie expuesta de dos grandes plataformas: el Banco Turks (formado por Grand Turk y pequeños cayos) y el Banco Caicos (South, East, Middle, West y North Caicos, Providenciales y algunos cayos), ocupando un área total aproximada de 900 km 2 (Fig. 1).
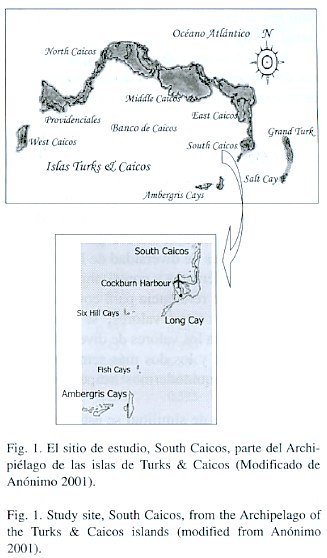
Trabajo de campo: el presente estudio se realizó durante el verano (julio-agosto) en las inmediaciones de South Caicos, la más pequeña de las islas mayores del Banco Caicos (Sealey 1994, Fig. 1). Se realizaron muestreos de crustáceos y equinodermos en ocho formaciones arrecifales comprendidas en los cayos: Long Cay (4), Big Ambergris Cay (2) y Fish Cay (2) y a cada formación arrecifal se le dio un nombre (se utilizan los nombres originales, en inglés, de las islas, cayos y sitios de muestreo). Los muestreos se llevaron al cabo con equipo de buceo autónomo y libre, en la zona de máxima profundidad de cada formación arrecifal. Se emplearon en cada sitio, en promedio, seis líneas de transecto de 15 m cada una, orientadas N-S y separadas 10 m. Se utilizaron seis cuadrantes de 1 m2 por transecto, separados cada 3 m, lo que dio un promedio de 36 cuadrantes por sitio de muestreo. Se realizó una prospección en cada sitio, con transectos y cuadrantes, donde se tomaron únicamente datos de presencia/ausencia de especies. Posteriormente se identificaron in situ y a nivel de especie siempre que fue posible, los crustáceos y equinodermos presentes en cada cuadrante, lo mismo que el número de individuos por especie. También se clasificó la condición del sitio de muestreo como de alto, bajo o nulo grado de sedimentación; ello, con estudios paralelos (de otro estudio e investigador, por lo que no se incluyen aquí) de comunidades de algas, donde se determinó la abundancia de algas asociadas a sedimentos, así como una estimación de la cantidad promedio de sedimentos para cada sitio de muestreo.
Análisis de datos: con los datos de presencia/ausencia se realizaron las curvas de especies/área (número de cuadrantes), para conocer el área mínima de muestreo, que sirvió de base para el muestreo subsecuente en cada sitio. Para analizar los patrones espaciales se calculó la media y la varianza del número de individuos por especie, con lo que se determinó si la distribución de los organismos en cada sitio era al azar, uniforme o agregada (Southwood 1978).
Con la finalidad de poder comparar los datos de número de especies de sitios con áreas de muestreo diferentes, se dividió el número de veces que se encontró cada especie en un cuadrante respecto al total de los cuadrantes (área) muestreados y se multiplicó por 100, para estandarizar a 100 m2 . Con los datos estandarizados para cada sitio se calcularon los índices de diversidad y equitabilidad de Shannon (Magurran 1988, Zar 1998). Para evaluar si los valores de los índices de diversidad de los sitios de estudio eran diferentes y con el objetivo de tener un punto de referencia para considerar diferencias entre estos valores, se realizaron pruebas de t para los valores de diversidad más alto y más bajo y los dos más semejantes, de crustáceos y equinodermos respectivamente (Zar 1998).
Los valores de similitud se estimaron con los datos de presencia/ausencia, por medio de comparaciones pareadas de cada especie en cada sitio. Se utilizó el índice de similitud de Bray-Curtis, que considera datos cualitativos de presencia/ausencia (Pielou 1984). Este índice es considerado el más adecuado para datos de comunidades bénticas (Bloom 1981, Magurran 1988). Se construyó una matriz de similitud y se clasificaron las comunidades de las formaciones arrecifales por medio de un dendograma derivado de la matriz de similitud (Southwood 1978).
Resultados
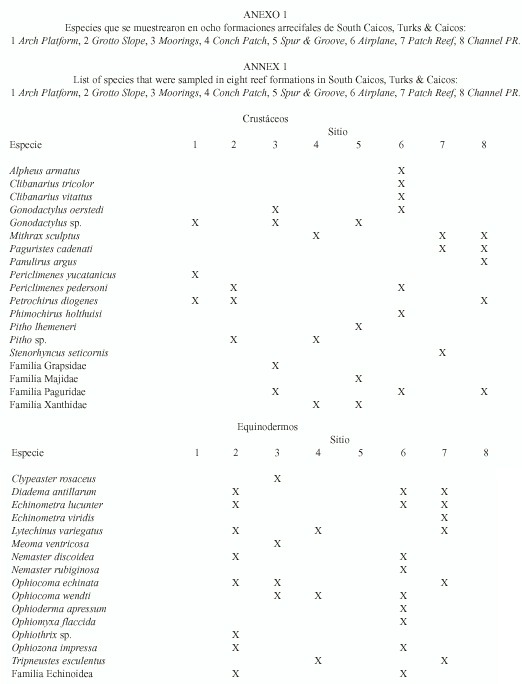
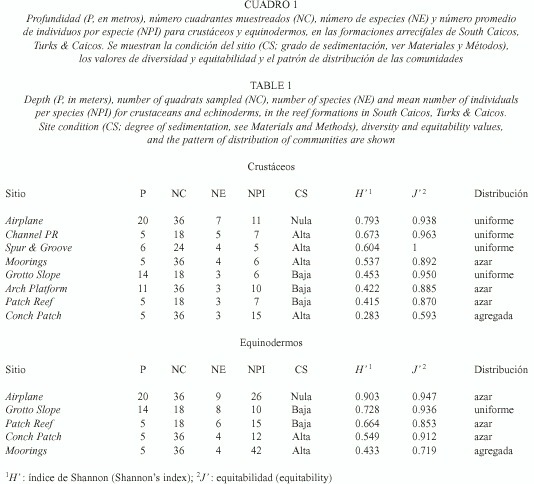
Diversidad: las ocho formaciones arrecifales estudiadas variaron en cuanto a profundidad (5 a 20 m), grado de sedimentación y especies muestreadas (Cuadro 1; la lista de especies se presenta en el Anexo). Únicamente en cinco de estas formaciones se encontraron equinodermos; por ello, los análisis se hicieron con base en cinco sitios para éstos y ocho para los crustáceos.
Una vez determinadas las curvas de especies/área, se estimó el área mínima de muestreo para cada uno de los ocho sitios, de manera que siempre se realizaron los muestreos un poco por encima del área mínima respectiva. El número de cuadrantes muestreados por sitio (18, 24 o 36 cuadrantes, determinado por la extensión del sitio), el número de especies (crustáceos 3 a 7 especies; equinodermos 4 a 9) y el número promedio de individuos por especie (crustáceos 5 a 15 individuos; equinodermos 10 a 42) variaron entre sitios (Cuadro 1).
Se encontraron diferencias significativas para los valores de diversidad para ambos grupos (crustáceos y equinodermos), tanto entre los valores extremos de los índices de diversidad (p < 0.01) como entre los más semejantes (p < 0.01). Para ambos grupos, el sitio con mayor diversidad fue Airplane que fue, a su vez, el único sitio que mostró nula sedimentación y donde la distribución espacial de crustáceos y equinodermos fue uniforme y al azar, respectivamente (Cuadro 1).
Respecto a los crustáceos, los siguientes tres sitios con valor más alto de diversidad fueron aquellos que presentaron alta sedimentación (Channel PR, Spur & Groove y Moorings; Cuadro 1). Por el contrario, para los equinodermos los tres sitios subsecuentes en diversidad fueron en los que se observó baja sedimentación (Grotto Slope, Patch Reef y Conch Patch; Cuadro 1). Asimismo, de los tres sitios donde no se encontraron equinodermos, dos presentaron alta sedimentación (Channel PR y Spur & Groove; Cuadro 1).
El único sitio con sedimentación alta donde se encontraron equinodermos (Moorings) presentó el valor más bajo de diversidad (Cuadro 1) y en donde, a pesar de que se muestrearon 48 individuos (la mayor abundancia encontrada de todos los sitios), 34 de éstos pertenecen a una sola especie (Clypeaster rosaceus). Este sitio, junto con el de menor diversidad para crustáceos (Conch Patch), presentaron los valores más bajos de equitabilidad y fueron los únicos con una distribución espacial agregada (Cuadro 1).
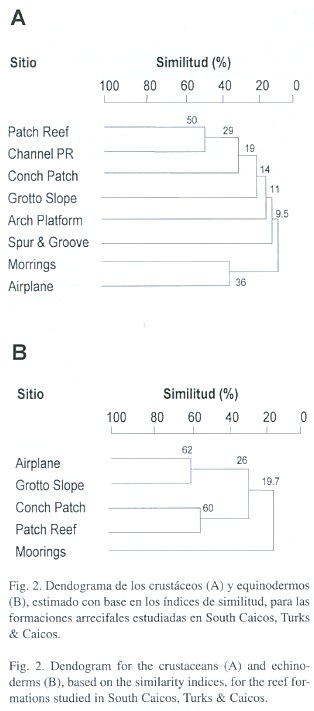
Similitud: para los crustáceos, los valores de similitud entre sitios fueron en general marcadamente bajos, con 13 comparaciones (de 28; 46%) que presentaron 0% de similitud. El valor más alto fue entre los sitios Patch Reef y Channel PR (50%). Se observa que sólo estos dos sitios forman un grupo que se separa, relativamente, del resto de los sitios (Fig. 2). Por el contrario, para los equinodermos se encontraron dos grupos (sitios) con porcentajes altos de similitud (Grotto Slope y Airplane: 62.5% y Conch Patch y Patch Reef: 60%), claramente reflejados en el dendograma (Fig. 2). El sitio de menor similitud (Moorings: 19.7%) fue el menos diverso y el único con sedimentación alta.
Discusión
La diversidad local de los arrecifes está relacionada con factores físicos que se correlacionan con la profundidad y las diferencias entre hábitats, mientras que la estructura ecológica está determinada principalmente por las interacciones entre organismos y las respuestas diferenciales a perturbaciones (Kleypass et al. 1999, Cornell y Karlson 2000, Vázquez-Domínguez 2000, Wood 2001). En este estudio se encontró que el sitio con mayor diversidad y alta equitabilidad para ambos grupos (Airplane), fue la única formación arrecifal donde se registró nula sedimentación y donde la comunidad presentó el mayor número de especies.
Aunado a lo anterior, cambios en parámetros como el número de especies o la abundancia relativa son un reflejo de cómo la sedimentación, como perturbación, puede afectar la estructura de la comunidad (Richmond 1993, Shinn et al. 2000). De tal forma que una comunidad bien desarrollada, con un alto número de parches de coral como se observa en Airplane, presenta cierta heterogeneidad ambiental –seguramente relacionada con la profundidad de este sitio– la que presumiblemente permite el establecimiento y desarrollo de un mayor número y variedad de organismos, en contraste con sitios con baja heterogeneidad y alta sedimentación (De Vantier et al. 1998, Cornell y Karlson 2000). Esto, a su vez, se refleja en los patrones de distribución observados en Airplane, donde los individuos ocupan todo el espacio disponible de una forma relativamente uniforme o al azar (Southwood 1978, Begon et al. 1996).
Nuestros resultados tienen limitaciones temporales y espaciales importantes, dado que los muestreos se realizaron en una sola época del año y sin réplicas; tampoco puede discriminarse (estadísticamente) entre los diferentes niveles de sedimentación ni de heterogeneidad ambiental entre sitios. Ello permite hacer únicamente descripciones generales y a escala local, que podrían variar en diferentes épocas o a lo largo del tiempo (Lirman y Biber 2000), sobre todo porque las poblaciones arrecifales están en un estado de flujo y cambio continuo, no sólo en una escala histórica (Jackson 1997, 2001), sino en tiempos ecológicos (Wood 2001).
Sin embargo, considerando dichas limitaciones y conociendo la biología de los organismos y sus requerimientos de hábitat, es posible describir de manera general la relación entre los parámetros poblacionales observados y la sedimentación (Holbrook et al. 2000, Zuschin et al. 2001). Después del sitio de mayor diversidad (Airplane), los siguientes sitios más diversos para los crustáceos presentaron alta sedimentación (Channel PR, Spur & Groove y Moorings). Esto puede estar relacionado con las características particulares de hábitat de las especies presentes: habitan en huecos en el sedimento (Gonodactylus), en conchas vacías que son abundantes en estos sitios con sedimento (Petrochirus) o en pastos marinos que se desarrollan en ambientes con alta sedimentación (Pitho, Mithrax). Sin embargo, otras especies quedan excluidas, como las que requieren de sustratos rocosos para establecerse (Alpheus) o que habitan entre los corales (Stenorhyncus, Periclimenes) (Sullivan 1991). Estos géneros, aunque poco abundantes en la zona de estudio en general, sí se encontraron en Airplane.
Por el contrario, para los equinodermos los sitios más diversos después de Airplane presentaron sedimentación baja (Grotto Slope y Patch Reef). El hecho de que la mayoría de las especies de equinodermos tienen requerimientos de hábitat que generalmente se presentan en sitios con poca o nula sedimentación, ya que habitan dentro de las esponjas, sobre y entre los corales o entre las rocas (Ophiocoma, Ophioderma, Ophiomyxa, Nemaster) (Sullivan 1991), pudo determinar los patrones observados. De acuerdo con lo anterior, en dos de los tres sitios donde no se encontraron equinodermos se registró alta sedimentación. El sitio con alta sedimentación donde sí se observaron equinodermos (Moorings), también fue el que presentó menor diversidad y donde más del 80 por ciento de los individuos muestreados fueron de la especie Clypeaster rosaceus, que habita exclusivamente en pastos marinos; nuevamente, de sitios con alta sedimentación. Asimismo, Moorings y Conch Patch presentaron el valor más bajo de equitabilidad, lo que significa cierta dominancia (Magurran 1988) y fueron los únicos donde se observó una distribución agregada. Ello puede asociarse con los patrones de distribución observados, ya que ciertos organismos se establecen en áreas particulares del ambiente donde tienen mayor probabilidad de sobrevivencia (i.e. parches menos perturbados) y otros, como el caso de C. rosaceus y Pitho sp., se distribuyen de manera agregada en manchones de pastos marinos (Sullivan 1991).
Recientemente se ha señalado que la transición de algunos arrecifes del Caribe de una estructura dominada por algas hacia una con baja densidad de algas y alta de corales, puede ser resultado de la presencia de Diadema antillarum (Edmunds y Carpenter 2001). De nuestros resultados, aunque sólo puede especularse al respecto, es interesante mencionar que en tres de los sitios con menor sedimentación (Grotto Slope, Airplane y Patch Reef) se encontraron individuos de D. antillarum. La relación entre la distribución y diversidad observadas y las características ambientales de los sitios se refleja también en los resultados de similitud. Para los equinodermos se obtuvieron dos grupos con altos valores de similitud. Uno formado por los dos sitios más diversos y de baja sedimentación, los cuales comparten cinco especies que habitan en rocas, entre los corales y, en general, en hábitat con poca sedimentación. El otro lo forman los sitios que siguen en diversidad y que comparten tres especies que se alimentan predominantemente de algas. Para los crustáceos se observó similitud entre los sitios Patch Reef y Channel PR, que aunque difieren en el grado de sedimentación se encuentran a la misma profundidad y en la misma zona, cercanos entre sí y además comparten dos especies (Mithrax y Paguristes), ambas de ambientes con cierta sedimentación. El resto de los sitios no forman grupos definidos como lo encontrado para los equinodermos, lo que sugiere que dichos sitios tienen otras características particulares que los hacen diferentes, donde pueden estar actuando otra serie de factores bióticos y abióticos (Sebens 1994).
Las perturbaciones –naturales o no– son un proceso importante para mantener la estructura y diversidad de las comunidades arrecifales, pero que varían en magnitud y cuyo efecto dependerá de la cronología y de las interacciones de los eventos de perturbación (Wood 2001). Por ello, detectar estos diferentes estados en los arrecifes requiere de estudios con escalas espacio-temporales amplias (Hughes 1994, Jackson 2001). Sin embargo, desde el punto de vista de conservación, es fundamental contar con información desde básica y general hasta detallada y de largo plazo, acerca de las características básicas y de la biología natural de las poblaciones y las comunidades. Los resultados observados en este estudio se refieren a diferentes grados de sedimentación natural (i.e. no asociada con actividades antropogénicas). Sin embargo, es importante hacer la distinción de que la sedimentación derivada de actividades humanas que puede afectar, entre otras, las características de resiliencia, persistencia y resistencia propias de la comunidad, no necesariamente tiene las mismas consecuencias (Richmond 1993, Vázquez-Domínguez et al. 1998, Nyström et al. 2000, Nyström y Folke 2001). A visa de comparación, una constante y alta sedimentación resultado del dragado, minado del coral y arena y, más importante, del mal manejo del terreno en las cuencas, afecta significativa y negativamente las comunidades arrecifales, en ocasiones con consecuencias letales (Maragos 1993, Shinn et al. 2000, Vázquez-Domínguez 2000). Así, la descripción de las características básicas de estructura poblacional y de historia natural de estas comunidades arrecifales en South Caicos, aunado al conocimiento de la naturaleza, distribución e impactos negativos de las actividades humanas, provee información útil para la planeación de futuras actividades de restauración y conservación de este ecosistema.
Agradecimientos
El trabajo de campo del presente estudio forma parte de la campaña realizada durante el curso Mapping Marine Ecosystems: Remote sensing and aerial photography techniques for mapping marine habitats, organizado por la Universidad de Boston y The School for Field Studies. Se da el más extensivo agradecimiento a los participantes del curso, grupo con el que la autora realizó el trabajo de campo, recabando información sobre diferentes aspectos, sitios y grupos taxonómicos, de los cuales este trabajo forma sólo una parte. Tres revisores anónimos hicieron valiosos comentarios con lo que el manuscrito es ahora sustancialmente mejor. La autora contó con una beca por parte de The School for Field Studies para participar en el curso.
Resumen
Se estimó la riqueza, diversidad, abundancia y distribución espacial de crustáceos y equinodermos en formaciones arrecifales de South Caicos, y se evaluó la relación entre las características de sedimentación en los arrecifes y dichos parámetros poblacionales. Estos arrecifes presentaron un gradiente de perturbación que va desde con un alto grado de sedimentación, cubiertos casi en su totalidad por comunidades de algas, hasta sin partículas de sedimentos depositados sobre los corales. Los sitios se clasificaron como de alta, baja o nula sedimentación y se estimó la riqueza, abundancia, diversidad, la distribución espacial de las especies y la similitud entre sitios. No se encontró un patrón único: tanto para crustáceos como equinodermos, el sitio con mayor diversidad y alta equitabilidad, presumiblemente asociada a la heterogeneidad ambiental que presenta esta formación arrecifal, también mostró nula sedimentación. Los dos sitios con menor diversidad para ambos grupos, aunque con baja sedimentación uno y alta el otro, presentaron el valor más bajo de equitabilidad y fueron los únicos con un patrón de distribución agregado. Para los crustáceos, los siguientes sitios en valor de diversidad fueron los de sedimentación alta, probablemente debido a que la mayoría de las especies presentes habitan comúnmente en huecos en el sedimento, conchas vacías o pastos marinos. Para los equinodermos, por el contrario, los siguientes sitios fueron los de sedimentación baja. Los requerimientos de hábitat de estas especies (dentro de esponjas, sobre corales o entre rocas), pudieron determinar este resultado. Los sitios con la diversidad más baja presentaron niveles altos de sedimentación. En estos sitios se encontró el valor más bajo de equitabilidad y una distribución espacial agregada para los crustáceos, mientras que la comunidad de equinodermos estaba dominada por una sola especie. Aunque los resultados observados sólo nos permiten hacer descripciones generales, el conocer las características poblacionales y de historia natural de estas comunidades arrecifales, aunado al conocimiento de las perturbaciones asociadas con las actividades humanas, provee información útil para la planeación de actividades de restauración y conservación de este ecosistema.
Referencias
Begon, M., J. Harper & C. Townsend. 1996. Community stability and community structure, p. 762-783. In M. Begon, J. Harper & C. Townsend (eds). Ecology: individuals, populations and communities Third Ed. Blackwell, Oxford, UK.
Bloom, S.A. 1981. Similarity indices in community studies: potential pitfalls. Mar. Ecol. Prog. Ser. 5: 125-128. [ Links ]
Connell, J.H. 1978. Diversity in tropical rainforests and coral reefs. Science 199: 1302-1310. [ Links ]
Cornell H.V. & R.H. Karlson. 2000. Coral species richness: ecological versus biogeographical influences. Coral Reefs 19: 37-49. [ Links ]
Chiappone, M., K.M. Sullivan & C. Lott. 1996. Hermatypic scleractinian corals of the southeastern Bahamas: a comparison to western Atlantic reef systems. Carib. J. Sci. 32: 1-13. [ Links ]
De Vantier, L.M., G. Death, T.J. Done & E. Turak. 1998. Ecological assessment of a complex natural system: a case study from the Great Barrier Reef. Ecol. Appl. 8: 480-496. [ Links ]
Edmunds, P.J. & R.C. Carpenter. 2001. Recovery of Diadema antillarum reduces macroalgal cover and increases abundance of juvenile corals on a Caribbean reef. Proc. Natl. Acad. Sci. USA 98: 5067-5071. [ Links ]
Ezcurra, E., A. Valiente-Banuet, O. Flores-Villela & E. Vázquez-Domínguez. 2001. Vulnerability to global environmental change in natural systems and rural areas: A question of latitude? pp. 217-246. In J.X. Kasperson, R.E. Kasperson (eds). Global environmental risk. United Nations University, Tokyo.
Gleason, D.F. 1998. Sedimentation and distributions of green and brown morphs of the Caribbean coral Porites astreoides Lamarck. J. Exp. Mar. Biol. Ecol. 230: 73-89.
Goreau, T.F., N.I. Goreau & T.J. Goreau. 1979. Corals and coral reefs. Sci. Am. 241: 728-734.
Holbrook S.J., G.E. Forrester & R.J. Schmitt. 2000. Spatial patterns in abundance of a damselfish reflect availability of suitable habitat. Oecologia 122: 109-120. [ Links ]
Hughes, T.P. 1994. Catastrophes, phase shifts, and largescale degradation of a Caribbean coral reef. Science 265: 1547-1551. [ Links ]
Huston, M.A. 1985. Patterns of species diversity on coral reefs. Annu. Rev. Ecol. Syst. 16: 149-177. [ Links ]
Jackson, J.B.C. 1997. Reefs since Columbus. Coral Reefs 16: S23-S32. [ Links ]
Jackson, J.B.C. 2001. What was natural in the coastal oceans? Proc. Natl. Acad. Sci. USA 98: 5411-5418. [ Links ]
Karlson, R.H. & H.V. Cornell. 1999. Integration of local and regional perspectives on the species richness of coral assemblages. Am. Zool. 39: 104-112.
Kleypas, J.A., J.W. McManus & L. A. Menez. 1999. Environmental limits to coral reef development: where do we draw the line? Am. Zool. 39: 146-159.
Lirman D. & P. Biber. 2000. Seasonal dynamics of macroalgal communities of the northern Florida reef tract. Bot. Mar. 43: 305-314. [ Links ]
Magurran, A.E. 1988. Ecological diversity and its measurement. Princeton University, Princeton, New Jersey. 192 p. [ Links ]
Maragos, J.E. 1993. Impact of coastal construction on coral reefs in the United-States-affiliated Pacific islands. Coastal Manag. 21: 235-269.
Nyström, M. & C. Folke. 2001. Spatial resilience of coral reefs. Ecosystems 4: 406-417. [ Links ]
Nyström, M., C. Folke & F. Moberg. 2000. Coral reef dis-turbance and resilience in a human-dominated environment. Trends Ecol. Evol. 15: 413-417.
Pielou, E.C. 1984. The interpretation of ecological data. Wiley, New York. 288 p.
Pointer, I.R. & R. Kennedy. 1984. Complex patterns of change in the macrobenthos of a large sandbank following dredging. Mar. Biol. 78: 335-352.
Reaka-Kudla, M.L. 1997. The global biodiversity of coral reefs: a comparison with rain forests, pp. 83-108. In M.L. Reaka-Kudla, D. E. Wilson & E. O. Wilson (eds). Biodiversity II: understanding and protecting our biological resources. National Academy Press. Washington, DC.
Richmond, R.H. 1993. Coral reefs: present problems and future concerns resulting from anthropogenic disturbance. Am. Zool. 33: 524-536. [ Links ]
Rogers, C.S. 1990. Responses of coral reefs and reef organisms to sedimentation. Mar. Ecol. Prog. Ser. 62. 185-202. [ Links ]
Sealey, N.E. 1994. Bahamian landscapes: an introduction to the geography of the Bahamas. Second ed. Media Publishing. Nassau, Bahamas. 115 p. [ Links ]
Sebens, K.P. 1994. Biodiversity of corals, what are we loosing? Am. Zool. 34: 115-133. [ Links ]
Shinn E.A., G. W. Smith, J.M. Prospero, P. Betzer, M. L. Hayes, V. Garrison & R.T. Barber. 2000. African dust and the demise of Caribbean coral reefs. Geophys. Res. Lett. 27: 3029-3032.
Southwood, T.R. 1978. Ecological methods with particular reference to the study of insects populations. Chapman & Hall, London. 524 p. [ Links ]
Sullivan, K.M. 1991. Guide to the shallow-water marine habitats and benthic invertebrates of the Exuma Cays Land and Sea Park, Bahamas. Bahamas National Trust. Sea and Sky Foundation, Coral Gables, Florida. 194 p.
Vázquez-Domínguez, E. 2000. La importancia de la biodiversidad arrecifal. The importance of reef biodiversity, p. 9-17. In O. Aburto Oropeza & C.A. Sánchez Ortíz (eds.). Recursos arrecifales del Golfo de California. Reef Resources of the Gulf of California. Universidad Autónoma de Baja California Sur, México.
Vázquez-Domínguez, E., J. González-Cano & V. Arenas. 1998. Áreas prioritarias marinas de uso de recursos, p. 127-150. In L. Arriaga Cabrera, E. Vázquez-Domínguez, J. González-Cano, R. Jiménez Rosenberg, E. Muñoz López & V. Aguilar Sierra (coords). Regiones prioritarias marinas de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, México.
Wood, R. 2001. Biodiversity and the history of reefs. Geol. J. 36: 251-263. [ Links ]
Wu, R.S. 1982. Periodic defaunation and recovery in a subtropical epibenthic community, in relation to organic pollution. J. Exp. Mar. Biol. Ecol. 64: 253-269. [ Links ]
Zar, J.H. 1998. Biostatistical analysis. Fourth Ed. Prentice-Hall, New Jersey. 718 p. [ Links ]
Zuschin M., J. Hohenegger & F.F. Steininger. 2001. Molluscan assemblages on coral reefs and associated hard substrata in the northern Red Sea. Coral Reefs 20: 107-116. [ Links ]
REFERENCIAS DE INTERNET
Anónimo, 2001. Information Directory for the Turks and Caicos Islands. TCI Mall, Cable & Wireless, Turks & Caicos Islands. Consultado: 10 julio 2001 http://www.tcimall.tc/turks/index.htm
1 Instituto de Ecología, UNAM. Ap. Postal 70-275, Ciudad Universitaria. México, DF 04510, México; evazquez@miranda.ecologia.unam.mx