











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkThe Baird's tapir (Tapirus bairdii) is one of four species of the genus Tapirus (Family Tapiridae; T. bairdii, T. pinchaque, T. terrestris and T. indicus), and the largest native terrestrial mammal in Neotropical ecosystems (Eisenberg, 1989; Emmons & Feer, 1990; Barongi, 1993). The ecosystem role of tapirs as largest herbivores include their potential role as seed dispersers (Rodrigues, Olmos & Galetti, 1993; Olmos, 1997; Fragoso, Silvius, & Correa, 2003; Tobler, Janovec & Conejo 2010; Capece, AliagaRossel & Jansen 2013; O'farrill, Galetti & Campos-Arceiz, 2013) or keystone browsers, maintaining disturbance of habitat heterogeneity in tropical forests (Williams & Petrides, 1980; Janzen 1981, 1982a, 1982b, 1983; Naranjo 1995a; Salas & Fuller, 1996; Talamoni & Assis, 2009; Chalukian, Bustos, & Lizarraga, 2013).
Once common in most vegetation types between sea level and subalpine paramo throughout Central America and northern South America, the Baird's tapir is now extinct in El Salvador and endangered throughout the rest of its range (Fragoso, 1991; March, 1994; IUCN, 2011). T. bairdii was classified as the tenth rarest Neotropical forest mammal by Dobson and Jinping (1993) based on population density and distribution. Habitat loss and overhunting are the principal causes for the decline in numbers (Emmons & Feer, 1990; Fragoso, 1991; Vaughan, 2011). They are an important food source for much of the rural population throughout their distribution (Eisenberg, 1989; Fragoso, 1991; Freese & Saavedra, 1991). Nevertheless, biological and ecological information is limited on Baird's tapir (Terwilliger, 1978; Naranjo, 1995a, 1995b; Alger, Vaughan & Foerster, 1998; Foerster, 1998; Valdez & Foerster, 2001; Foerster & Vaughan, 2002).
It is known that Baird's tapir consumed more than 100 plant species in different vegetation types of tropical dry and humid forests of Costa Rica and Panama (Terwilliger, 1978; Janzen, 1982a; Williams, 1984; and Naranjo, 1995a). Terwilliger (1978) reported that Baird's tapirs on Barro Colorado Island, Panama spent approximately 88% of their active time feeding. Because of the limited detailed seasonal behavioral data on diet and foraging, the goal of this study was to describe the seasonal diet and foraging behavior of a female Baird's tapir in a Costa Rican lowland rainforest.
Methods
Study area: This study was conducted in Corcovado National Park (CNP) (41,789 ha, 8:264 to 8:394 N and 83:254 to 83:454 W) located on the southwestern coast of the Osa Peninsula, Costa Rica. The mean annual temperature in CNP fluctuates between 23 and 26.5 C, depending on altitude. The rainy season extends from May to November, and the dry season from December to April (Vaughan 1981). Created in 1975 (Vaughan, 1981, 2012), CNP represents the largest remaining tract of Pacific lowland tropical rainforest in Central America and supports an estimated 500 tree species, 367 bird species, 140 mammals, 51 amphibians and 64 reptiles (Ross, 2013, Boza, 1992; and Hartshorn, 1991). Naranjo (1995b) estimated a population of between 155 and 249 Baird's tapirs in CNP.
The tapir was studied around the Sirena ranger station, located on the southwest coast of CNP. The regions north and northwest of Sirena were alluvial, well drained soils where past agricultural practices resulted in a mosaic of primary and secondary forests in varying degree of regeneration (Phillips, 1993). East and southeast of the station was dominated by primary forests on steep slopes. Two major rivers provided drainage in the region: the Rio Claro to the south of the Sirena station and the Rio Sirena to the northwest. Numerous intermittent streams were found throughout the study area.
Data collection: Tapir 132 was located in the field using a Telonics TR-2 receiver and RA-14 antenna (Telonics Inc., Meza, Az. 85204). At five minute intervals, data was collected on diet and foraging activity. Because of the relative tranquil nature of tapir 132, we had a unique opportunity to collect field data. No other tapir 132 in the study area allowed us to collect data as did tapir; therefore the study is limited to one individual.
Tapir 132 was observed for a minimum of 20 hrs. per month from June 1995 to April 1996, excluding August 1995. Considering that tapir activity was predominantly nocturnal (Foerster & Vaughan, 2002), diet and foraging behavior data was collected between 1800-0600 hrs. Observation periods lasted between two and 11 hrs.
Diet: At five minute intervals, the following data was recorded for the first food consumed: a) plant part eaten (leaf, fruit bark, stem, or flower), (b) processing time of bite, and (c) plant identification number. Initially, a sample of each plant species eaten was mounted, pressed and assigned a reference number. In subsequent observations, plants were either identified by their reference number or a sample was collected for comparison with those in the collection.
Foraging activity: State of activity was recorded at five minute intervals and classified as one of the following: a) feeding - in the act of taking a bite, b) standing chewing - stationary and chewing, b) moving chewing - walking or running while chewing, c) standing not chewing - stationary, not chewing, d) moving not chewing - running or walking, not chewing, and e) social interaction - actions directed towards or affected by another tapir. A continuous record was kept of behaviors such as defecation, urination, drinking, scent marking, intra/interspecies interactions and vocalizations.
Bites and paces: The total number of bites taken and number of paces traveled between each bite was recorded during the first ten minutes of each half hour. A hand held audio recorder was used to record field observations.
Biomass consumed: Data on biomass (gms. dry wt.) consumed was gathered in December, January and February. Samples of the same plant species approximating the amount taken for the first bite taken during five minute intervals were collected and dried in the sun to a constant weight. An average bite weight was calculated and multiplied by the average number of bites taken per ten minute period to calculate total biomass consumed per unit of time.
Data analysis
The replicated G test (William's correction) was used to compare ratio of major food items to total items eaten, a) ratio of major food items to total items eaten in wet season vs dry season, b) wet season vs study long ratios, c) dry season against study long ratios, d) monthly ratios of food items against study long ratios of the category, e) ratios of activity states vs total observations, f) monthly ratios of feeding activity vs study long ratios. The Pearson Correlation Coefficient was used to test the degree of association among diet and foraging behaviors (Sokal & Rohlf, 1995).
Results Diet
Plant species in diet: Tapir 132 was observed for 286 hrs. of which she was active for 181 hrs. Average duration of observation periods was 4.4 hrs. (n=65). She consumed 126 different plant species of which 35,2% were vines, 34,1% trees, 15.9% shrubs, and 14.8% herbs. Three species accounted for 40% of her diet for the entire study. Monstera sp., a vine in the Areaceae Family was the most common species consumed in all months except July and constituted 23,1% of her diet. Although consumed in only three of the ten months of the study, the fruit of Persea sp. (Lauraceae family) accounted for 9.9% her diet. Psychotria sp., a shrub in the Rubiaceae family, represented the third most common plant eaten (7,0%).
Percent composition by major food items: Study long totals show the Tapir 132 diet consisted of 67.0% leaf matter, 18,6% fruit, 11,7% stem, 2,1% bark and 0,1% flower (n=2172, Digital Appendix 1). Wet season totals were 66,9% leaf, 22,3% fruit, 7,9% stem, and 3,0% bark (n=1389). Dry season diet consisted of 68,1% leaf, 12,4% fruit, 18,8% stem, 0,4% bark and 0,3% flower (n=783).
Percent fruit and bark in the diet were significantly higher in the wet season than in the dry season (Gadj=54,1692, P<0.001; Gadj=51,5773, P<0,001 respectively) while percent stem in the diet was significantly lower in the wet season than in the dry season (Gadj= 126,6221, P<0,001, Digital Appendix 2). Similarly, wet and dry season percentages of fruit, stem and bark were significantly different than study long totals for the respective categories (Digital Appendix 2).
The June diet for Tapir 132 consisted mainly of leaf (65,5%) and fruit (20,1%) while stem and bark were minor components (Figure 1). June was the only month during the study where bark formed a notable percentage of the diet (7,5%). All bites registered for bark were of a small tree species, Psychotria sp. (Rubiaceae Family).
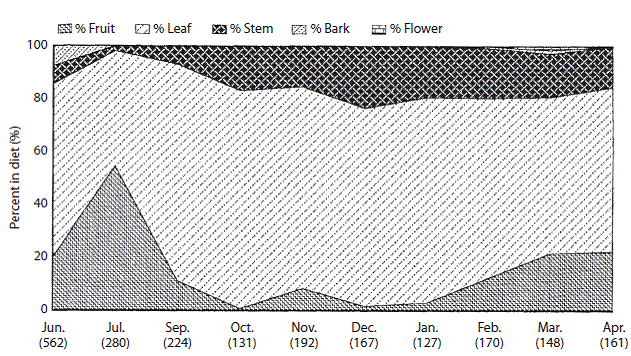
Figure 1: Percent composition of major food items consumed by tapir 132 from June 1995 to April 1996. Numbers in parenthesis are monthly sample sizes.
In July, fruit replaced leaf as the highest percentage in the diet (54,3%). The sharp increase in fruit intake was due primarily to the consumption of Persea sp. (Lauraceae Family). This species accounted for 44,3% of the total July diet and 81,6% of the fruit consumed in July. Stem matter was inconsequential during this period, while bark and flower were absent from the diet.
September through January (wet season) diets were relatively constant, with leaf as the most common food type eaten, followed by stem and fruit. No bites were recorded for either bark or flower during this interval. February and March (dry season) showed a slight decrease of leaf and stem in the diet, replaced by an increase of fruit. Bark and flower were infrequently registered in this period. April values of percent leaf, fruit and stem remained relatively stable from March values. Neither bark nor stem were recorded during this period.
The ratios of leaf to total observations in July and March were significantly lower than the study long leaf ratio (Gadj=62,82; P<0,001; Gadj=4,40; P<0,05 respectively) (Digital Appendix 3). September through January ratios of leaf to total observations were significantly higher than the study long leaf ratio. A significantly higher ratio of fruit to total observations was consumed in July compared to the study long ratio (Gadj=172,52; P<0,0001). September through February ratios of fruit to total observation were significantly lower than the study fruit long ratio. The ratios of stem to total observations in June, July and September were significantly lower than the study long stem ratio. December through February ratios of stem to total observations were significantly higher than the study long stem ratio (Digital Appendix 3). The ratio of bark to total observations in June was significantly higher than the study long ratio (Gadj=7,35; P<0,01) (Digital Appendix 3). It was the only month during the study when bark had a significant amount of bark consumption.
Foraging behavior
Feeding (F): Average percent time spent feeding (F) for the entire study period was 29,2% Percent feeding observations for wet and dry seasons were 28,3 and 1,3% respectively. The wet season ratio of feeding to total observations were significantly higher than the dry season ratio (Gadj=4,6800, P<0,05; Digital Appendix 4). Nevertheless, neither the wet or dry season feeding ratios were significantly different than the study long ratio. Monthly ratios of feeding to total observations did not vary significantly over time (Digital Appendix 5). Study long peaks were observed in July (23,2%) (low) and September (33,3%) (high) (Figure 2).
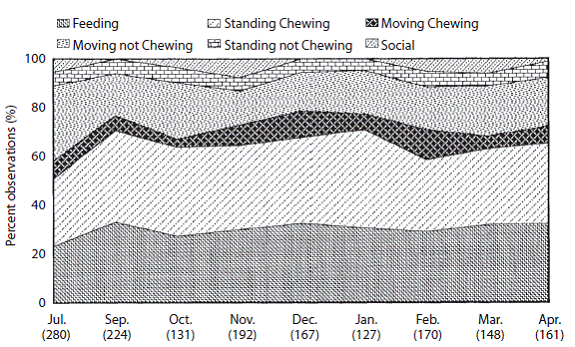
Figure 2: Percent observations of different activity states for tapir 132 from July 1995 to April 1996. Numbers in parenthesis are monthly sample sizes.
Standing chewing (SC): Average percent time spent standing chewing (SC) over the entire study period was 33,4% (Digital Appendix 5). Percent SC observations for wet and dry seasons were 33.4 and 33,5% respectively. Neither the wet or dry season ratios of SC to total observations were significantly different than the study long ratio (Digital Appendix 4). Similarly, wet and dry seasons ratios did not differ. Only the July ratio of SC to total observations was significantly different than the study long ratio (Gadj=4,15, P<0,05)(Digital Appendix 6). Study long peaks were observed in January (40,3%) (high) and February (29,6) (low) (Digital Appendix 5).
Moving chewing (MC): Average percent time spent moving chewing (MC) for the entire study period was 7,6% (Digital Appendix 5). Percent MC observations for wet and dry season were 6,7 and 8,7% respectively. The wet season ratio of MC to total observations was significantly higher than the dry season ratio (Gadj=5,6125; P<0,025, Digital Appendix 4). Nevertheless, neither the wet or dry season ratios of MC observations were significantly different than the study long ratio. Monthly ratios of MC to total observations varied significantly over time (Gadj=14,24; P<0,01, Digital Appendix 6). October and February ratios of MC were significantly different than the study long ratio (Gadj=4,57, P<0,05; Gadj=5,01; P<0,05 respectively)(Digital Appendix 6). Study long peaks were observed in October (3,4%)(low) and February (12,5%)(high).
Standing not chewing (SNC): Average percent time spent standing not chewing (SNC) for the entire study period was 5,7% (Digital Appendix 5). Percent SNC observations for wet and dry season were 5,7 and 5,8% respectively. Neither the wet or dry season ratios of SNC to total observations were significantly different than the study long ratio (Digital Appendix 3). Similarly, the wet season ratio was not different than the dry season ratio.
Moving not chewing (MNC): Average percent time spent moving not chewing (MNC) for the entire study period was 20,1% (Digital Appendix 5). Percent MNC observations for wet and dry season were 21,8 and 18,3% respectively. The wet season ratio of MNC to total observations was significantly lower than the dry season ratio (Gadj=7,7545; P<0,01, Digital Appendix 4). Nevertheless, neither wet nor dry season ratios of MNC were significantly different than the study long ratio. Study long peaks were observed in July (30,6%)(high) and November (14,0%)(low)( Digital Appendix 5).
Social interactions (Si): Average percent time spent in social interactions (SI) for the entire study period was 3.4% . Percent SI observations for wet and dry season were 4,2 and 2,6% respectively (Digital Appendix 5). The wet season ratio of SI to total observations was significantly lower than the dry season ratio (Gadj=8,4266; P<0,005, Digital Appendix 4). Nevertheless, neither wet nor dry season ratios of SI were significantly different than the study long ratio.
Paces taken during 10 minute periods: The study long average for number of paces taken during ten minute periods was 42,9 (n=147, s.d.=46,36). Multiplied by her average stride length (1,02 m, n=50), it is estimated that Tapir 132 traveled 262.5 m per hour. The wet and dry season averages for paces per ten minute periods were 50,2 (s.d.=54,95) and 34,8 (32,68) respectively. The wet season values for paces per ten minute interval were significantly greater than in the dry season (F=4,11, d.f. =1, 145, P=0,0461). Study long peaks were observed in July (91,8) (high) and December (23,1) (low)(Figure 3).
Bites taken during ten minute periods: The study long average for number of bites taken during ten minute periods was 49,7 (n=147, s.d.=30,14). The wet and dry season averages for bites taken during ten minute periods were 42,1 (s.d.=29,21) and 58.2 (s.d.=29,07) respectively. The wet season values for bites per ten minute interval were significantly smaller than dry season paces per ten minute period (F=11,18, d.f.=1, 145; P=0,0011).
Study long peaks occurred in July (20,2) (low) and January (70,5) (high) (Figure 3).
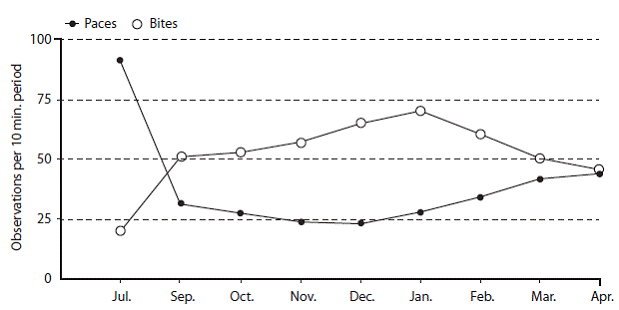
Correlations
Correlations among foraging behaviors: Negative correlations were observed between percent feeding observations and percent MNC observations and paces taken during ten minute periods (Digital Appendix 7). There was a negative association between percent observations of SC and paces per 10 min period and a positive correlation between SC observations and bites per 10 min period. Percent observations of MNC was positively correlated with paces per 10 min period and negatively associated with bites per 10 min period. A strong negative correlation was observed between the number of bites and paces taken per 10 min period (Digital Appendix 7).
Correlation of diet to foraging activities: Percent fruit in the diet was strongly associated with number of paces traveled (positive correlation), and the mean number of bites (negative correlation, Digital Appendix 8). Higher percent fruit in the diet was moderately associated with lower percent observations of SC and MNC (positive).
Correlation of percent leaf matter, paces and bites: A strong negative correlation was calculated between percent leaf matter and number of paces. Percent leaf in the diet was moderately, positively associated with both percent SC observations and number of bites taken. A weak, negative association was shown between percent leaf matter and observations of MNC. A strong, positive correlation was observed between percent stem and the number of bites taken. A moderate, negative correlation was observed between percent stem and of paces traveled.
Plant biomass consumed
Biomass consumed: Average biomass consumed per bite (dry wt.) calculated over a three month period was 3,32 gms (n=417). Multiplied by the three month average number of bites taken during ten minute intervals (49,7) and the number of active hours per day (12), it is estimated that Tapir 132 consumed an average of 11.9 kilos per day or 4,307 kilos of biomass (dry wt) per year. The average bite weight (dry wt.) for December was 3.95 gms. (n=155). In January her grams per bite were calculated at 3,35 gms. (n=105 bites) and for February, 2,67 grams per bite (n=157).
Discussion Diet and seasonality
Baird's tapir consumed more than 100 plant species in different vegetation types of tropical dry and humid forests of Costa Rica and Panama (Terwilliger, 1978; Janzen, 1982a; Williams, 1984; and Naranjo, 1995a). In the present study, Tapir 132 consumed 126 different plant species. Williams and Petrides (1980) found similar results studying T. indicus in Malaysia, reporting more than 115 species consumed. Chalukian et al (2013) found T. terrestris fed on 57 species of plants based on fecal samples in El Rey National Park in Argentina.
Using fecal analysis and direct observations, Naranjo (1995a) reported the diet of Baird's tapir in CNP as 65,4% leaf matter, 25,2% fiber, and 9,4 % fruit from at least 94 species and 36 plant families. According to Naranjo (1995a), tapirs consumed more fruit in the dry season while fiber was more frequently ingested in the wet season. The study long total for percent leaf in the diet of Tapir 132 was similar to Naranjo's (1995a) results (65,4 vs. 67,0% in this study) and it was similar in the wet and dry seasons. However, percent fruit was much higher for tapir (9,4 vs. 18,6% in this study) and fruit was consumed in greater quantities in the wet season, contrary to Naranjo's (1995a) results. These differences may be partly explained by examining the data points associated with the fruit of Persea sp. The fruit of Persea sp. is relatively large (13 cm x 9 cm) with a correspondingly large seed and a soft fleshy mesocarp. Tapir 132, as well as other tapirs observed, consumed only the mesocarp of the fruit. It is doubtful that any remains would be recognizable after passing through the digestive system of a tapir and would explain the absence of evidence for this species in fecal samples examined by Naranjo (1995a). Within Tapir 132's home range, the distribution of Persea sp. is restricted to an area of approximately 30 hectares of primary forest northwest of the Sirena ranger station (Foerster & Vaughan, 2002). Finally, the Persea sp. seeds were observed on the ground only in wet season months and accounted for 66,6% of Tapir 132's fruit intake in the wet season. Removing Persea sp. data points from this study decreases percent fruit in Tapir 132's diet from 18,6% to 9,2% for the year, almost equal to Naranjo's (1995a) results. Also, seasonal values become more equivalent to Naranjo's (1995a) results and percent fruit intake would be higher in the dry season than the wet season. This behavior is probably common to other tapirs in the Sirena region. During every observation period in which Tapir 132 visited the Persea sp. patch, at least one other tapir was spotted feeding on the same fruit. On one occasion, four tapirs were observed feeding on Persea sp. under the same tree, but fruits were left. Bodmer (1990) analyzed stomach contents and fecal samples for T. terrestris, finding that this species consumed 33% fruit and 67% leaf and stem, similar to the present study and Naranjo's (1995a) work in the leaf consumption. Including stem confuses the interpretation.
Given the estimated consumption of about 11,9 kilos of biomass (dry wt) per day or 4,307 kilos per year. Naranjo (1995b) estimated between 155-249 tapirs in CNP. If the total tapir population in CNP was calculated as 200 tapirs, the total biomass consumed by tapirs in the park would be 872,400 kilos of biomass (dry wt) yearly, which could greatly impact the park's vegetation, especially those species consumed.
Foraging Behaviors
Terwilliger (1978) reported that Baird's tapirs on Barro Colorado Island, Panama spent approximately 88% of their active time feeding. Similarly, the majority of Tapir 132's active hours were dedicated directly to foraging. By combining totals for observations of the F, SC and MC activity states, 71% of awake time was spent feeding on or processing some food item. In only 25 of the 2172 data points did Tapir 132 pass the entire five minute observation period without taking a bite, of which 15 could be attributed to either a social encounter or when she became inactive within the observation periods. Generally speaking, more than a minute rarely passed that she did not take a bite.
In Peru, Bodmer (1990) estimated lowland tapir (T. terrestris) diet by stomach, caecal and fecal samples as 33% fruit and 67% leaf and fiber. Bodmer's results suggest that T. terrestris feeds on a fruit occurring in monotypic clumps and thus maximizes the benefits of frugivory by minimizing searching costs. Optimal foraging theories predict that large non-ruminant herbivores should resort to frugivory only if the nutritional gains outweigh the searching costs (Stephens and Krebs 1986). Tapir 132 may have exhibited this strategy in those months when Persea sp. was available. Although this species does not occur in strictly monotypic stands, Tapir 132 would feed almost exclusively on Persea sp. for two to three hours every night when available.
Several correlations of percent fruit in the diet to foraging behaviors also support this seasonal frugivory hypothesis. Increased fruit in the diet was correlated with an increase in observations of MNC, reflecting the travel time between fruiting trees. Accordingly, the number of paces taken per unit of time also increased during this time. A strong negative correlation was seen when values for the three foraging behavior categories in which Tapir 132 was actually eating (F, SC and MNC) were combined. This is confirmed by a similar decrease in the number of bites taken per unit of time. This is logical when you consider that the average handling time for Persea sp. was 15 times greater than that for other food types.
In other words, Tapir 132 was willing to travel farther and take fewer bites to increase her fruit consumption.
Given that bark and flower were minor components of Tapir 132's diet in most months of the study, variations in monthly percent of fruit in the diet were primarily reflected in negative correlations with percent leaf matter. As a result, where percent fruit was positively associated with MNC observations and paces taken per unit of time, percent leaf was negatively correlated with those same categories. Where percent fruit was negatively correlated with SC observations and number of bites per ten minute periods, positive associations were seen with percent leaf consumption. This is the only tapir study combining diet and foraging, therefore it is impossible to compare results and correlations with other research.
Ecosystem role of tapirs as keystone browsers and seed dispersers
Tapir 132 was found to consume an average of 11.9 kilos of biomass (dry wt) per day or 4,307 kilos per year. Naranjo (1995b) estimated between 155-249 tapirs in CNP. If the total tapir population in CNP was calculated as 200 tapirs, the total biomass consumed by tapirs in the park would be 872,400 kilos of biomass (dry wt) yearly. If this was converted to wet biomass, over a million kilos of biomass would be consumed by tapirs in CNP. The results of this study and Naranjo (1995a) make Baird's tapir one of the largest terrestrial neotropical herbivores/ keystone browsers and important seed dispersers in CNP. Extrapolating, all tapir species fulfill important roles within neotropical terrestrial ecosystems (Fragoso 1997; Fragoso & Huffman 2000; Henry, Feer & Sabatier 2000; Barcelos, Bobrowiec, Santoiotti & Gribel 2013; O'farrill, Galetti & Campos-Arceiz 2013: Tobler, Janovec & Conejo 2010). Dirzo and Miranda (1990) consider the tapir a potentially important indicator species for the health of neotropical rainforests. For these reasons and the abundant T. bairdii population in certain areas of CNP, protection, research and conservation efforts should continue.

