











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Pleopeltis Humb. & Bonpl. ex Willd. es uno de los géneros más diversos de helechos dentro la familia Polypodiaceae con aproximadamente unas 90 especies, principalmente representadas en el neotrópico. Es un género en constante revisión taxonómica y ha estado sujeto a modificaciones en busca de la formación de grupos naturales (Smith et al., 2006; Prado & Hirai, 2010; Smith &Tejero-Díez, 2014; PPG I, 2016). Las apomorfías más importantes en el género son las escamas peltadas persistentes y los nectarios foliares; aunque estos caracteres pueden estar ausentes en algunas especies (Otto, Janßen, Kreier, & Schneider, 2009; Smith &Tejero-Díez, 2014). En Colombia el género Pleopeltis está representado por aproximadamente 20 especies según la nueva circunscripción de Smith y Tejero-Díez (2014). Pleopeltis macrocarpa (Bory ex.Willd.) Kaulf. es una especie epifita, de rizomas largamente rastreros, hojas monomorfas ampliamente espaciadas a lo largo del rizoma, con escamas peltadas, ovaladas o lanceoladas, soros impresos circulares u ovalados de hasta 6 mm de diámetro con escamas receptaculares peltadas usualmente clatradas que cubren todo el soro hasta la madurez, etapa en la que se caen. Las esporas son monoletes con ornamentación verrugosa (Hennipman, Veldhoen, & Kramer, 1990). P. macrocarpa es una especie que se distribuye en todo el neotrópico, África, Madagascar y la India. En Colombia es de amplia distribución, pero está mejor representada en la región andina, Sierra Nevada de Santa Marta y Serranía de la Macarena y se encuentra hasta los 3 500 m de altitud (Murillo, Murillo, & León, 2016).
En relación a la esporogénesis en monilophyta se registran trabajos detallados que describen a nivel estructural y ultraestructural la esporogénesis en varios grupos de helechos eusporangiados. Parkinson (1987) describe la esporogénesis de Psilotum nudum (L.) P. Beauv. resaltando la importancia del tapete y su estrecha relación con la ontogenia de las esporas y el esporodermo. Brown y Lemmon (2001a) describen en detalle los aspectos ultraestructurales de la esporogénesis en Angiopteris Adans. (Marattiales) concluyendo que esta especie presenta meiosis monoplastídica. Por el contrario, a la meiosis poliplastidial que describieron durante la esporogénesis de Ophioglossum L. (Ophioglossales) (Brown & Lemmon 2001b). Por otro lado, Rincón, Forero, Gélvez, Torres y Rolleri (2011) describen el desarrollo del estróbilo y la esporogénesis en Equisetum giganteum L. de ejemplares recolectados en Colombia. En Equisetum bogotense Kunth se ha descrito a nivel ultraestructural los eventos de la esporogénesis, el desarrollo del esporodermo y eláteres, siendo esta, una de las investigaciones que aporta descripciones detalladas sobre los procesos de la esporogénesis de Equisetales (Rincón, Torres, & Rolleri, 2013).
A pesar de que los helechos leptosporangiados son considerablemente más diversos con respecto a los eusporangiados (PPG I, 2016; Sessa, 2018). Los estudios sobre la esporogénesis y ontogenia de los esporangios son escasos, y solo algunos abordan aspectos ultraestructurales e histoquímicos que permiten entender los procesos fisiológicos que ocurren durante el desarrollo y maduración de los esporangios. En este sentido es notable mencionar los trabajos pioneros de Wilson (1958) y Pal y Pal (1963) quienes describen en detalle la ontogenia de los esporangios de Phlebodium aureum (L.) J. Sm. y Ceratopteris thalictroides (L.) Brongniart, respectivamente. Aunque son algunos de los trabajos más completos para su época y que marcan las pautas para posteriores investigaciones en ontogenia esporangial; en estos estudios no presentan evidencias fotográficas de sus observaciones. Posteriormente, algunas investigaciones incluyen observaciones e imágenes con microscopía electrónica de barrido (MEB) y de trasmisión (MET), así como la aplicación de novedosos métodos de inclusión y preparación de muestras con lo cual las descripciones del desarrollo del soro y los esporangios se tornaron precisos y minuciosos. Con estos avances se incursionó por primera vez en la descripción de eventos sub-celulares de la esporogénesis y ontogenia de los esporangios (Peterson & Kott, 1974; Bowen & Williams, 1977; Sheffield & Bell, 1979; Sheffield, Laird, & Bell, 1983; Lugardon, 1990; Van Uffelen, 1990, 1992, 1993; Parkinson, 1995; Qiu, White, & Turner, 1995; Churchill, Tryon, & Barrington, 1998; González, Prada, & Rolleri, 2010; Gabarayeva, Grigorjeva, & Márquez, 2011; Triana-Moreno, 2012).
En relación a aspectos morfo-anatómicos, se han hecho esfuerzos para esclarecer cuáles son las características que determinan que varias especies del género Pleopeltis sean consideradas tolerantes a la sequía y conocidas como los helechos de la resurrección, dada la capacidad para hidratarse rápidamente, después de permanecer secos y en latencia por largos períodos de tiempo, sin experimentar daño celular o histológico apreciable (Helseth & Fischer, 2005; Layton et al., 2010; John & Hasenstein, 2018). En este sentido toman especial relevancia los aspectos bioquímicos, y anatómicos de las raíces y los rizomas, pero en especial, con la estructura y composición de las paredes celulares de las escamas peltadas presentes en las regiones vegetativas de estas plantas (John & Hasenstein, 2017; Lagoria et al., 2018). A pesar de la importancia de las escamas peltadas como estructuras de protección de los receptáculos de Pleopeltis, los cuales cubren por completo a los soros durante toda su ontogenia, hasta la fecha, no se cuenta con estudios ontogenéticos y morfo-anatómicos de estos órganos que indique su papel en la protección e hidratación de los esporangios.
En esta investigación, se tiene por objetivo hacer un estudio detallado sobre la ontogenia de los esporangios y las escamas receptaculares presentes en los soros de P. macrocarpa, utilizando técnicas y equipos modernos. Se incluyen aspectos morfogenéticos por primera vez explorados y se resalta su relación con compuestos como la lignina, celulosa, pectinas, polifenoles totales, carbohidratos de reserva y estructurales y proteínas. Se analiza y se discute el desarrollo y la histoquímica de las escamas receptaculares y su importancia en los procesos de protección de los esporangios durante las etapas más lábiles de su desarrollo, así como los procesos de hidratación después de un período de sequía. Finalmente, se analiza la ornamentación de las esporas maduras y los procesos de formación y depósito del esporodermo.
Materiales y métodos
En marzo y mayo (época lluviosa) 2017 se recolectaron para su estudio más de 30 ejemplares fértiles de P. macrocarpa que crecen epífitos sobre los troncos de árboles en el vivero el Edén de las flores, Las Palmas km 3 antes del Aeropuerto José María Córdoba, municipio de Rionegro-Antioquia a una altitud aproximada de 2 260 m (6°09'58.73" N & 75°27'14.38" W). El material de referencia se depositó en el Herbario del Instituto de Biología de la Universidad de Antioquia (HUA) (Rincón 021).
Para los estudios de la ontogenia de los soros se tomaron 30 frondes fértiles con estructuras reproductivas o esporangios en diferentes etapas de desarrollo. Las cuales se fijaron en Glutaraldehído al 2.5 % en buffer fosfato 0.2 M pH 7.2 durante 24-48 h a 6 °C. Luego de la fijación, las muestras se lavaron en el mismo buffer seguido de agua destilada para posteriormente post-fijarlas con Tetróxido de Osmio al 2 % por 4 h a 6 °C en oscuridad y agitación constante. Posteriormente, se deshidrataron durante una hora en una serie gradual de etanol y durante 12 h etanol al 100 % a 6 °C. Las muestras se embebieron en mezclas progresivas de óxido de propileno-resina Spurr, por una semana a temperatura ambiente. Se hicieron varios cambios de resina pura durante seis días en agitación constante. La resina fue polimerizada a 60 °C por 48 h. Se obtuvieron secciones de 0.4-0.5 μm de grosor con cuchillas de vidrio en un Ultramicrótomo Leica®; las cuales se tiñeron con azul de Toluidina (TBO) en bórax al 1 %, pH 3.6 por 30-60 segundos. Muestras adicionales de las frondas se fijaron en una mezcla de formol, etanol y ácido acético (FAA) por 24-48 h a 6 °C, se cortaron en fragmentos de 1 cm de longitud, se deshidrataron en una serie gradual de etanol y xilol (Ruzin, 1999) y se incluyeron en Paraplast plus (Mc Cormick®) durante 12 h a 55 °C. Se obtuvieron secciones transversales con micrótomo rotatorio LEICA RM 2125, entre 5-7 μm de grosor. Finalmente, se procedió a la tinción de las secciones con los protocolos para descripción de tejidos en general y las pruebas histoquímicas específicas que se indican en la tabla 1 (Ruzin, 1999; Demarco 2014, 2017; Soukup, 2014; Yeung, Stasolla, Sumner, & Huang, 2015; Monteiro & Demarco, 2017).
TABLA 1 TABLE 1 Tinciones y pruebas histoquímicas aplicadas a las secciones de los soros de Pleopeltis macrocarpa Stains and histochemical tests applied to the sori sections of Pleopeltis macrocarpa
| TINCIÓN | COMPUESTO/ESTRUCTURA QUE RESALTA |
| Azul de Toluidina (TBO) | Paredes secundarias lignificadas (xilema y esclerénquima), se tiñen de color verde o azul verdoso. Paredes primarias con polisacáridos carboxilados y ácido péctico (parénquima, colénquima, floema y cambium), se tiñen de color purpura o purpura rojizo. Polifenoles se tiñen de azul grisáceo. Esporopolenina de azul turquesa. |
| Safranina-Azul de alcian (Fasga) | Paredes primarias se tiñen de azul. Paredes secundarias, cutícula, núcleos, suberina y polifenoles se tiñen de color rojo o fucsia. |
| Azul de Alcian-Hematoxilina de Gill III | Resalta paredes celulares primarias en azul, núcleos en color violeta oscuro, polifenoles marrón oscuro y lignina marrón claro. |
| Técnica de Wiesner (Fluoroglucinol-HCL) | Específico para lignina (grupos coniferil aldehído y sinapil aldehido), se resalta en color rojo. La tinción no es permanente. No se observan reacciones cruzadas con otros compuestos o estructuras. |
| Fast Blue B | Polifenoles se observan de color rojo a marrón oscuro. Es específico y no presenta reacciones cruzadas. |
| PAS-Amidoblack | Polisacáridos (de reserva y estructurales) se observan con coloración fucsia. Proteínas y núcleos se tiñen de color azul intenso. Presenta reacción cruzada con polifenoles de color marrón. |
| Rojo de Rutenio | Específico para sustancias pécticas de paredes primarias, las cuales se observan de color rojo o fucsia. Presenta reacciones cruzadas con los núcleos celulares. |
| Calcofluor White M2R | Paredes celulósicas microfibrilares (coloración azul). Se observa con epifluorescencia a longitudes de onda de excitación de 340-380 nm (luz UV) y emisión 420 nm (azul). |
Para llevar a cabo las descripciones morfológicas con MEB, el material se fijó en Glutaraldehído como se indicó anteriormente y se deshidrató en 2.2 dimetoxipropano (Lin, Falk, & Stocking, 1977) para posteriormente desecar a punto crítico con un desecador SAMDRI®-795. Los fragmentos de las frondas con los soros se montaron sobre cinta conductiva de carbono de doble cara y se recubrieron con oro en una ionizadora DENTON VACUUM DESK IV durante 5 min. Las observaciones y registro de las imágenes se realizaron en un microscopio electrónico de barrido JEOL JSM-6490LV.
Las secciones en resina y parafina se examinaron con un microscopio fotónico Nikon 80i eclipse® y Nikon eclipse Ni con el sistema de contraste diferencial de interferencia (CDI). Además, las secciones en parafina también fueron analizadas con epifluorescencia, empleando el filtro de triple banda de excitación DAPI-FITC-Texas el cual incorpora un filtro de excitación con ventanas de paso de banda angostas en las regiones espectrales violeta (395 a 410 nm), azul (490 a 505 nm) y verde (560 a 580 nm). Las fotografías se obtuvieron con cámara digital Nikon DS-Fi1® utilizando el programa NIS Elements versión 4.30.02 de Nikon.
Los términos empleados para las descripciones se usaron sensuLellinger (2002) y Punt, Hoen, Blackmore, Nilsson y Le Thomas (2007). El término banda de orgánulos se utiliza según Brown y Lemmon (2001a), el de cámaras plasmodiales según Rincón et al. (2011) y orbículas, según Tryon y Lugardon (1991) y Passarelli, Gabriel, Prada y Rolleri (2010). El término escamas receptaculares sensuHennipman et al. (1990) es utilizado como sinónimo de paráfisis escamosas peltadas.
Resultados
Las frondas fértiles de P. macrocarpa se caracterizan por presentar soros impresos exindusiados, circulares u ovalados de hasta 6 mm de diámetro, y que, por lo general, forman una fila, cada una localizada submarginalmente sobre la cara abaxial de la fronda (Fig. 1A). Los esporangios son los típicos leptosporangios de pedicelos largos con dos filas de células, de anillo en posición vertical de aproximadamente 16 células y un estomio bien desarrollado (Fig. 1B). En las condiciones de epifluorescencia (filtro de triple banda de excitación DAPI-FITC-Texas) más Calcofluor el anillo de los esporangios presenta una fuerte autofluorescencia en el amarillo-naranja, indicando la presencia de lignina y posiblemente suberina. Las células de pared del esporangio y parte del pedicelo se aprecian de color azul, por la presencia de celulosa; las esporas presentan autofluorescencia en el amarillo indicando la presencia de esporopolenina (Fig. 1B).
Los soros son de desarrollo mixto, es decir, en un mismo soro hay esporangios en diferentes etapas del desarrollo, son superficiales y asociados a tejido vascular (Fig. 1C, Fig. 1D; Fig. 5C). Al llegar a la madurez, los esporangios se encuentran recubiertos por completo por las escamas receptaculares (Fig. 1E), momento en el cual, éstas se desprenden del soro (Fig. 1F, Fig. 1G). El soro inicia su desarrollo en el receptáculo, éste se observa como una depresión en la cara abaxial de la fronde. Las células epidérmicas del receptáculo se caracterizan por ser alargadas de núcleos prominentes y paredes primarias delgadas (Fig. 1H). Estas células del receptáculo por divisiones mitóticas darán origen a los esporangios y las escamas receptaculares. En el primer caso, las células epidérmicas se dividen por un plano oblicuo dando origen a la célula basal y apical, que posteriormente se diferenciarán en el primordio del esporangio (Fig. 2A). Por el contrario, las células epidérmicas que formarán las escamas receptaculares se dividen en un plano periclinal no oblicuo.
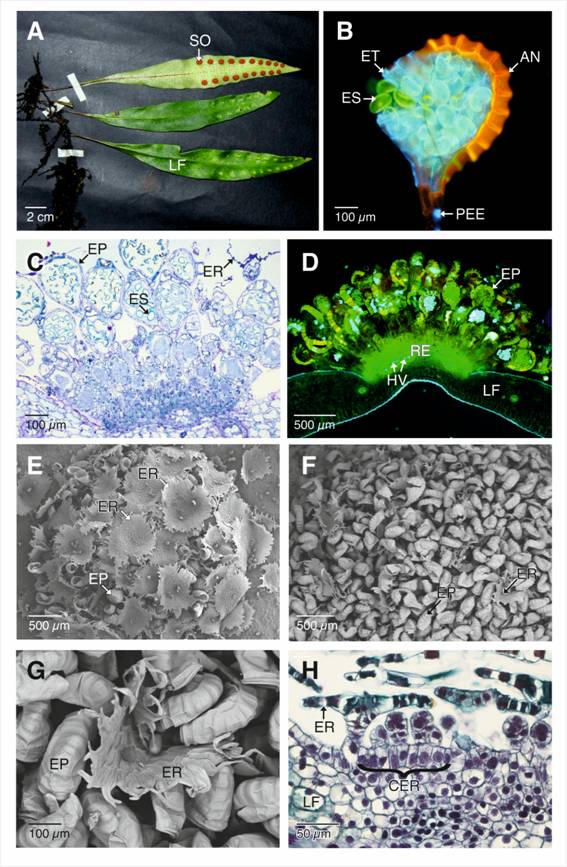
Fig. 1. Fig. 1 Soros, esporangios maduros y células receptaculares de Pleopeltis macrocarpa. A. Frondas fértiles. B. Montaje en fresco y sin fijar del esporangio maduro y esporas (Calcofluor y epifluorescencia con filtro DAPI-FITC-Texas). C. Sección longitudinal de los soros maduros (azul de Toluidina). D. Sección longitudinal a mano alzada de los soros maduros (sin teñir, epifluorescencia con DAPI-FITC-Texas). Se aprecian esporangios en diferentes etapas del desarrollo y la autofluorescencia de los anillos de los esporangios, indicando la presencia de lignina y probablemente suberina. E-G. Soros maduros. E. Soro cubierto por escamas receptaculares. F-G. Escamas receptaculares desprendiéndose del soro. H. Características citológicas de las células epidérmicas receptaculares. AN: anillo del esporangio; CER: células epidérmicas del receptáculo; EP: esporangios; ER: escamas receptaculares; ES: esporas; ET: estomio; LF: lamina foliar; HV: haces vasculares; PEE: pedicelo de los esporangios; RE: receptáculo; SO: soros. Sori, mature sporangia and receptacle cells of Pleopeltis macrocarpa. A. Fertile fronds. B. Fresh mount and unfixed of the mature sporangia and spores (Calcofluor and epifluorescence with a DAPI-FITC-Texas filter). C. Longitudinal section of the mature sori (Toluidine blue O). D. Freehand longitudinal section of the mature sori (unstained, epifluorescence with DAPI-FITC-Texas filter). Sporangia are seen at different stages of development, annulus autofluorescence that indicate the presence of lignin and probably suberin. E-G. Mature sori. E. Sorus covered by receptacle scales. F-G. Receptacle scales detaching from the sorus. H. Cytological features of the epidermal cells of the receptacle.
AN: sporangium annulus; CER: epidermal cells of the receptacle; EP: sporangia; ER: receptacle scales; SP: spores; ET: stomium; LF: leaf blade; HV: vascular bundles; PEE: stalks of the sporangia; R: receptacle; SO: sori.
Ontogenia de las escamas receptaculares: Las células epidérmicas que darán origen a las escamas receptaculares experimentan, al inicio del desarrollo, divisiones en el plano periclinal, dando origen a una columna multicelular uniseriada, ligeramente inclinada, con una evidente célula apical convexa (Fig. 2B). Posteriormente, una célula de contorno deltoide, ubicada en el primer tercio basal de esta columna multicelular, se divide en un plano oblicuo formando dos células hijas, una dorsal y otra basal (Fig. 2C). Posteriores divisiones mitóticas de estas dos células dan origen a la parte proximal del escudo y a la región distal o alas, que, junto con el pedicelo, conforman las escamas receptaculares (Fig. 2D, Fig. 2E). Las células de los pedicelos de las escamas receptaculares y las células de la región proximal del escudo presentan núcleos en posición parietal y grandes vacuolas con depósitos de polifenoles (Fig. 2D). En contraste, las células de la región distal o alas del escudo, al llegar a la madurez se colapsan, carecen de núcleos y muestran depósitos de polifenoles (Fig. 2E). Al final del desarrollo, las escamas receptaculares están formadas por un largo pedicelo multiseriado, que es perpendicular al receptáculo y un escudo paralelo al mismo (Fig. 2E).
Ontogenia de los esporangios y esporogénesis: Una vez formadas la célula basal y apical experimentan divisiones mitóticas seriadas en diferentes planos para formar una estructura alargada y una porción expandida terminal globosa que corresponde al pedicelo y primordio de la cápsula del esporangio, respectivamente (Fig. 2B). A medida que se desarrollan los primordios de los esporangios, los esporocitos premeióticos terminan rodeados por un grupo de células parietales estériles a partir de las cuales, por divisiones mitóticas anticlinales y periclinales se formará la cápsula del esporangio (Fig. 2F, Fig. 2G). Los esporocitos premeióticos se dividen mitóticamente para formar una capa de células tapetales iniciales (Fig. 2H), las cuales se vuelven a dividir anticlinal y periclinalmente para formar un tapete biestratificado (Fig. 3A, Fig. 3B).
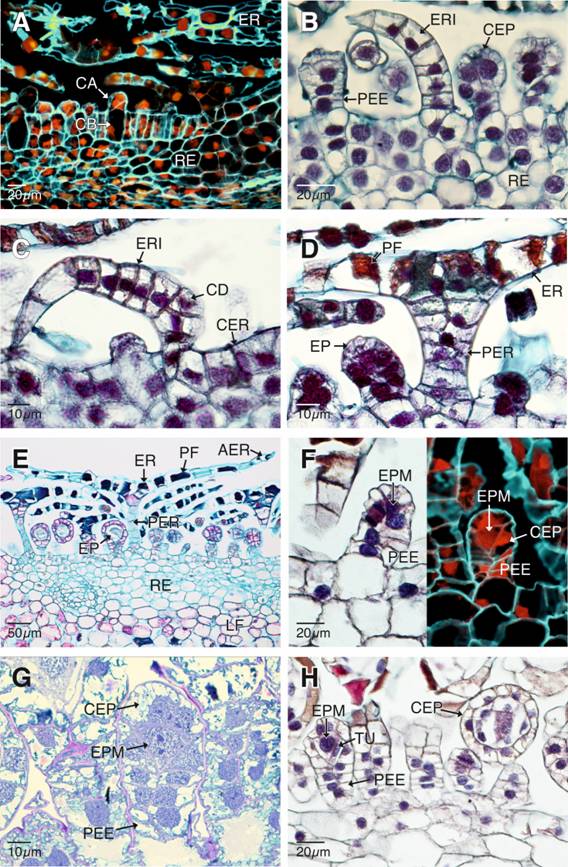
Fig. 2. Fig. 2 Ontogenia de los esporangios y escamas receptaculares de Pleopeltis macrocarpa. A. Divisiones mitóticas de las células epidérmicas del receptáculo (epifluorescencia con filtro DAPI-FITC-Texas). B. Esporangios y escamas receptaculares inmaduros. C-E. Escamas receptaculares en diferentes etapas de desarrollo. F-H. Etapas tempranas del desarrollo de los esporangios. F-G. Pedicelos y cápsulas de los esporangios en formación. Los esporocitos premeióticos se aprecian rodeados por una capa de células parietales (campo claro con epifluorescencia y filtro DAPI-FITC-Texas). H. Diferenciación de la capa uniestratificada del tapete tipo celular. AER: alas de las escamas receptaculares; CA: célula apical; CB: célula basal; CER: células epidérmicas del receptáculo; CD: célula de contorno deltoide; CEP: cápsulas de los esporangios; EP: esporangios; EPM: esporocitos premeióticos; ER: escamas receptaculares maduras; ERI: escamas receptaculares inmaduras; LF: lámina foliar; PEE: pedicelos de los esporangios; PF: polifenoles; PER: pedicelos de las escamas receptaculares; RE: receptáculo; TU: tapete celular de un estrato. Ontogeny of the sporangia and receptacle scales of Pleopeltis macrocarpa. A. Mitotic divisions of the epidermal cells of the receptacle (epifluorescence with a DAPIFITC-Texas filter). B. Sporangia and receptacle scales, both of them inmature. C-E. Receptacle scales on different development stages. F-H. Early stages of the sporangial development. F-G. Stalks and capsules of sporangia in formation. The premeiotical sporocytes are visible, surrounded by a layer of parietal cells (Bright-field with epifluorescence with a DAPIFITC-Texas filter). H. Differentiation of the single layered cellular tapetum.
AER: wings of the receptacle scales; CA: apical cell; CB: basal cell; CER: epidermal cells of the receptacle; CD: deltoid contoured cell; CEP: capsules of the sporangia; EP: sporangia; EPM: premeiotic sporocytes; ER: mature receptacle scales; ERI: immature receptacle scales; LF: leaf blade; PEE: stalks of the sporangia; PF: polyphenols; PER; stalks of the receptacle scales; RE: receptacle; TU: single layered cellular tapetum.
Para este momento del desarrollo, el tapete biestratificado delimita a cuatro esporocitos premeióticos, estos se caracterizan por presentar citoplasma escaso de aspecto granular, un núcleo central voluminoso, uno o dos nucléolos y cromosomas condensados (Fig. 3B). Los esporocitos premeióticos se dividen mitóticamente para aumentar el número de células que experimentarán la meiosis (Fig. 3C). A medida que el esporangio madura, el estrato interno del tapete pierde la integridad histológica y forma un plasmodio conocido como tapete plasmodial, que invade la cavidad esporangial rodeando a los esporocitos premeióticos y separándolos entre sí, mientras que el estrato externo del tapete persiste hasta la maduración de las esporas, finalmente degenera y desaparece para la liberación de las mismas (Fig. 3D). La invasión del plasmodio tapetal acelera la meiosis de los esporocitos, ahora, profásicos, que aumentan de tamaño y se tornan esféricos, de núcleos voluminosos, algo excéntricos muy granulares y vacuolados (Fig. 3D). En esta etapa del desarrollo se aprecian las cámaras plasmodiales que corresponden a espacios claros entre el plasmodio y los esporocitos. La membrana nuclear de los esporocitos en metafase I se desintegra por completo, los cromosomas se hacen más evidentes, mientras se observan masas de orgánulos desplazados periféricamente rodeando la zona donde se localizan los cromosomas, y paquetes de microtúbulos que participan en el proceso de segregación cromosómica (Fig. 3E). Así mismo, para esta etapa del desarrollo, las cámaras plamodiales se hacen evidentes y se observan gran cantidad de orgánulos y núcleos dentro del plasmodio. Al final de la meiosis II se aprecia una zona granular que separa los núcleos de la tétrada de esporas en formación. Esta zona corresponde a la banda de orgánulos que determinará el depósito de la placa celular que se sintetizará de manera simultánea separando a las cuatro esporas inmaduras que se disponen principalmente de forma decusada o tetragonal (Fig. 3F, Fig. 3G). Las tétradas de esporas inicialmente se encuentran juntas en la misma cámara plasmodial, pero progresivamente se separan hasta quedar en cámaras plasmodiales individuales, a partir de las cuales se alcanza la etapa de madurez y posterior liberación (Fig. 3H; Fig. 4A, Fig. 4B, Fig. 4C).
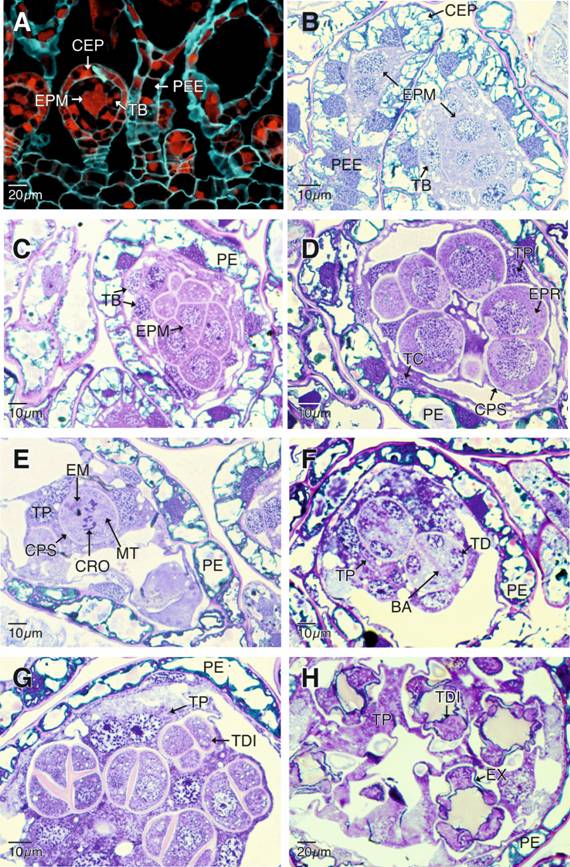
Fig. 3. Fig. 3 Desarrollo de los esporangios de Pleopeltis macrocarpa. A-B. Diferenciación del tapete biestratificado (epifluorescencia con filtro DAPI-FITC-Texas) y divisiones mitóticas de los esporocitos premeióticos (campo claro). C. Se observa al tapete biestratificado. En contacto con los esporocitos premeióticos que se dividen aun por mitosis. D. Esporocitos en profase I. E. Esporocitos en metafase I. F. Esporocitos al final de la meiosis II. G-H. Tétradas de esporas en disposición decusadas y tetragonales en diferentes etapas del desarrollo. BA: banda de orgánulos; CEP: cápsula del esporangio; CPS: cámaras plasmodiales; CRO: cromosomas; EM: esporocitos en metafase I; EPM: esporocitos premeióticos; EPR: esporocitos en profase I; EX: exosporio; MT: microtúbulos; PE: pared del esporangio; PEE: pedicelo del esporangio; TB tapete celular biestratificado; TC: estrato externo del tapete celular; TD: tétrada; TDI: tétradas inmaduras; TP: tapete plasmodial. Development of the sporangia of Pleopeltis macrocarpa. A-B. Differentiation of the bistratified tapetum (epifluorescence with a DAPI-FITC-Texas filter) and premeiotic sporocytes in mitotic divisions (Bright-field). C. The bistratified tapetum can be seen in contact with the premeiotic sporocytes still dividing by mitosis. D. Sporocytes in prophase I. E. Sporocytes in metaphase I. F. Sporocytes at the end of meiosis II. G-H. Tetragonal spores tetrads and decussately disposed during different development stages.
BA: band of organules; CEP: capsule of the sporangia; CPS: plasmodial gaps; CRO: chromosomes; EM: sporocytes on metaphase I; EPM: premeiotic sporocytes: EPR: sporocytes on prophase I; EX: exospore; MT: microtubules; PE: sporangium wall; PEE: pedicel of the sporangia; TB: bistratified cellular tapetum; TC: external strata of the cellular tapetum; TD: tetrad; TDI: immature tetrads; TP: plasmodial tapetum.
Las células que conformarán el anillo del esporangio se diferencian, las paredes periclinales y anticlinal interna se engrosan inicialmente por depósitos de materiales de pared primaria y en seguida, por depósitos de lignina, con lo cual se desarrolla el engrosamiento en forma de “U” (Fig. 4D). Las células del anillo inicialmente presentan una vacuola que ocupa la mayor parte del citoplasma, lo que relega al núcleo a una posición parietal. Durante el desarrollo, en estas vacuolas se depositan materiales polifenólicos (Fig. 4C). En este momento del desarrollo se observan a las esporas en sus cámaras plasmodiales individuales. Se aprecia claramente la disminución del tapete plasmodial y la presencia de estructuras esféricas u orbículas en el mismo (Fig. 4B, Fig. 4C).
La síntesis de la pared de la espora o esporodermo se da en el siguiente patrón: primero se deposita el exosporio, seguido del endosporio y finalmente el perisporio (Fig. 4A, Fig. 4B, Fig. 4C, Fig. 4D, Fig. 4E, Fig. 4F). El exosporio es la capa más gruesa del esporodermo y está formada por esporopolenina, le sigue en grosor el endosporio formado de materiales de pared primaria (Fig. 4C). El perisporio es la capa más delgada, también conformada de materiales de pared primaria, posiblemente algo de celulosa y pectinas. En algunas muestras analizadas el perisporio se observa algo desprendido del exosporio al momento de la liberación de las esporas (Fig. 4E, Fig. 4F). Las esporas maduras son monoletes con lesura engrosada, de ornamentación verrugosa con pequeñas orbículas esféricas distribuidas azarosamente sobre el esporodermo (Fig. 4G, Fig. 4H).
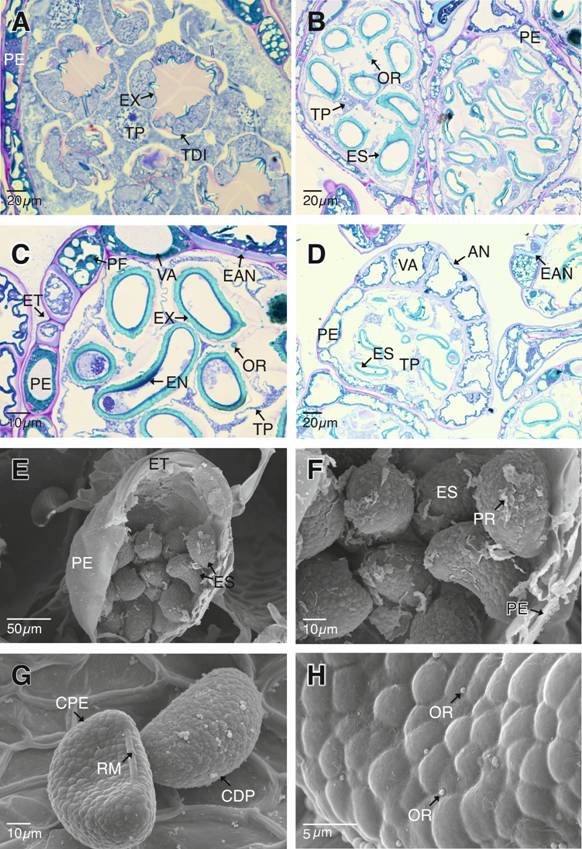
Fig. 4. Fig. 4 Tétradas de esporas, anillo del esporangio y esporas maduras de Pleopeltis macrocarpa . A. Tétradas de esporas en disposición tetragonal. B-C. Esporas en sus cámaras plasmodiales individuales. Endosporio y el exosporio se han diferenciado. C-D. Diferenciación de las células del anillo del esporangio. E-F. Estomio abierto y liberación de las esporas, las cuales presentan restos de perisporio. G-H. Esporas monoletes maduras con ornamentación verrugosa y orbículas sobre el esporodermo. AN: anillo de los esporangios; CDP: cara distal de las esporas; CPE: cara proximal de las esporas; EAN: engrosamientos en forma de “U” de las células del anillo del esporangio; EN: endosporio; ES: esporas; ET: estomio; EX: exosporio; RM: lesura monolete; OR: orbículas; PE: pared del esporangio; PF: polifenoles; PR: perisporio; TDI: tétradas inmaduras; TP: tapete plasmodial; VA: vacuola. Tetrads of spores, sporangium annulus and mature spores of Pleopeltis macrocarpa. A. Spore tetrads tetragonal disposed. B-C. Spores in their individual plasmodial gaps. The endospore and the exospore have differentiated. C-D. Differentiation of the annulus cells of the sporangium. E-F. Open stomium and release of the spores, which bear perispore remnants. G-H. Mature monolete spores with verrucate sculpture and orbicules over the sporoderm.
AN: sporangium annulus; CDP: distal face of the spores; CPE: proximal face of the spores; EAN: U shaped thickening of the sporangium annulus cells; EN: endospore; ES: spores; ET: stomium; EX: exospore; RM: monolete laesura; OR: orbicules; PE: sporangium wall; PF: polyphenols; PR: perispore; TDI: immature tetrads; TP: plasmodial tapetum; VA: vacuole.
Tinciones y pruebas histoquímicas: En etapas tempranas del desarrollo, las paredes celulares que conforman los esporangios y las escamas receptaculares están constituidas principalmente de celulosa y materiales de pared primaria (Fig. 5A). Sin embargo, a medida que maduran las células de los escudos de las escamas receptaculares pierden la afinidad por el Calcofluor, lo que se evidencia por la pérdida de fluorescencia azul y por el contrario, presenta autoflorescencia en el amarillo-naranja indicando el depósito de una cutícula gruesa (Fig. 5B).
Una vez que los esporangios y las escamas receptaculares alcanzan la madurez, solo los pedicelos de ambas estructuras y las células de la cápsula del esporangio presentan afinidad por el Calcoflúor y por consiguiente fluorescencia en el azul indicando la presencia de celulosa (Fig. 5C). En contraste, los anillos de los esporangios presentan autofluorescencia en el amarillo-naranja debido a la presencia de lignina y probablemente suberina.
Las pruebas para lignina indican la presencia de esta biomolécula únicamente en el anillo y el estomio de los esporangios maduros, así como en las células más próximas de los pedicelos a la cápsula del esporangio (Fig. 5D, Fig. 5E).
Durante los estadíos iniciales del desarrollo, los escudos de las escamas receptaculares están constituidas por células de paredes primarias engrosadas de citoplasma funcional (Fig. 5F, Fig. 5G). A medida que las escamas receptaculares alcanzan la madurez, las paredes de las células de los escudos se engrosan y las del centro del escudo se tornan pigmentadas (Fig. 5H).
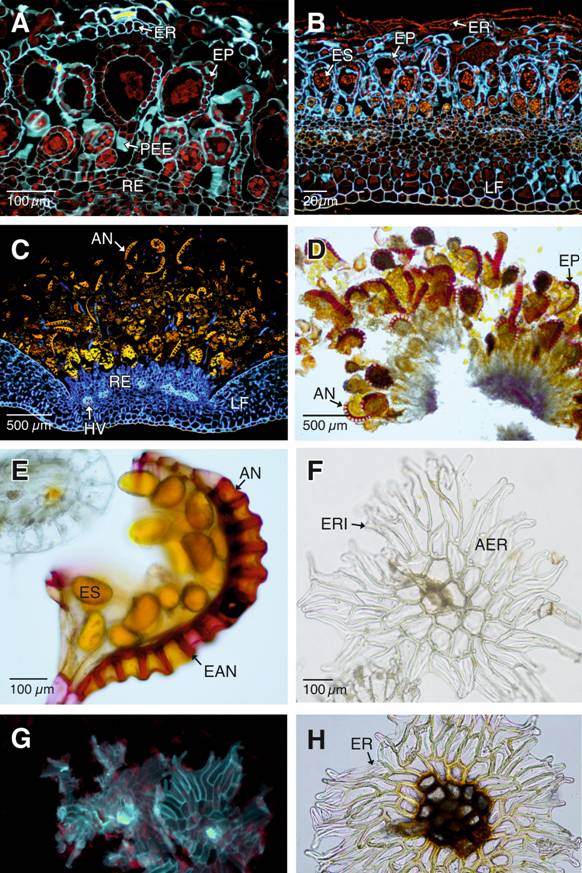
Fig. 5. Fig. 5 Pruebas histoquímicas de la ontogenia de los esporangios y escamas receptaculares. A-C. Esporangios y escamas receptaculares en diferentes momentos del desarrollo (Calcofluor y epifluorescencia con filtro DAPI-FITC-Texas). A. Sección longitudinal del soro inmaduro. Se aprecia la afinidad de los esporangios y escamas por el Calcofluor. B. Siguiente etapa de maduración del soro. Los escudos de las escamas receptaculares excluyen al Calcofluor. C. Soro con esporangios maduros (Calcofluor con epifluorescencia y filtro DAPI-FITC-Texas). D-E. Células del anillo y pedicelos de los esporangios con depósitos de lignina, en color rojo-fucsia (Fluoroglucinol ácido). F. Escama receptácular inmadura (sin tinción y campo claro) G. Escama receptácular inmadura con paredes celulares engrosadas por compuestos de pared primaria (Calcofluor y epifluorescencia con filtro DAPI-FITC-Texas). H. Escama receptácular madura (campo claro). AN: anillo de los esporangios; AER: alas de las escamas receptaculares; EAN: engrosamientos en forma de “U” de las células del anillo del esporangio¸ EP: esporangio; ER: escamas receptaculares maduras; ERI: escamas receptaculares inmaduras; ES: esporas; LF: lamina foliar; HV: haces vasculares; PE: pared del esporangio; PEE: pedicelo de los esporangios; RE: receptáculo. Histochemical tests during the ontogeny of the sporangial and receptacle scales. A-B. Longitudinal sections of the sori showing the mixed development of the sporangia and receptacle scales (Calcofluor and epifluorescence with a DAPI-FITC- Texas filter). A. Longitudinal section of the immature sorus. The affinity of the sporangia and scales towards Calcofluor can be observed. B. The next stage of the sorus maturation. The shields of the receptacle scales exclude Calcofluor. C. Sorus with mature sporangia (Calcofluor and epifluorescence with a DAPIFITC-Texas filter). D-E. Lignin deposits in cells of the annulus and stalks of the sporangia staining in red-fuchsia (acid phloroglucinol) F. Immature receptacle scale (unstained and Bright-field). G. Immature receptacle scale with cell walls thickened by primary wall constituent (Calcofluor and epifluorescence with a DAPI-FITC-Texas filter). H. Mature receptacle scale (Bright-field).
AN: sporangium annulus; AER: wings of the receptacle scales; EAN: U shaped thickening of the sporangium annulus cells; EP: sporangia; ER: mature receptacle scales; ERI: immature receptacle scales; ES: spores; LF: leaf blade; HV: vascular bundles; PE: sporangium wall; PEE: stalk of the sporangia; R: receptacle.
Estas paredes celulares dan negativo para lignina y adicionalmente pierden la afinidad por el Calcofluor y presentan autofluorescencia en el amarillo-naranja por la gruesa cutícula que presentan (Fig. 6A). En tanto que las alas de los escudos de las escamas receptaculares se aprecian colapsadas, sin núcleos, y con paredes engrosadas ricas en materiales de pared primaria (Fig. 6B) y materiales pectídicos (Fig. 6C, Fig. 6D) determinadas por la tinción de Toluidina y prueba específica del rojo de Rutenio, respectivamente.
En los soros, los polifenoles son muy abundantes desde las etapas tempranas del desarrollo de los esporangios y de las escamas receptaculares. Son especialmente abundantes en las vacuolas de las células maduras de las escamas receptaculares y en las células de los anillos y pedicelos de los esporangios (Fig. 6E, Fig. 6F).
Los almidones se aprecian como pequeños gránulos de color púrpura, son abundantes en las células epidérmicas, tejido parenquimatoso del receptáculo y en los primordios de los esporangios. En la madurez se detectaron en las células del pedicelo, y pared del esporangio, así como en el tapete bi-estratificado (Fig. 6G, Fig. 6H). Las proteínas no se apreciaron con claridad con esta coloración, excepto por la tinción azul intensa de los núcleos.

Fig. 6. Fig. 6 Aspectos histoquímicos y tinciones específicas de la ontogenia de los esporangios y escamas receptaculares. A. Escama receptaculares madura con paredes engrosadas (Calcofluor y epifluorescencia con filtro DAPI-FITC-Texas). B. Sección longitudinal de las alas de las escamas receptaculares maduras (Azul de Toluidina O). C-D. Escamas receptaculares maduras con cutículas engrosadas (Rojo de Rutenio). E. Esporangios y escamas receptaculares con polifenoles (Fast Blue B). F. Sección del anillo del esporangio con grandes vacuolas con depósitos de polifenoles (Azul de toluidina O). G-H. Carbohidratos de reserva y estructurales teñidos de color fucsia-rojo y proteínas en color azul oscuro (PAS-Amidoblack). AER: alas de las escamas receptaculares; AN; anillo de los esporangios; EP: esporangio; ER: escamas receptaculares maduras; ES: esporas; GA: gránulos de almidón; LF: lamina foliar; PF: polifenoles; PEE: pedicelo de los esporangios; PER: pedicelo de las escamas receptaculares; RE: receptáculo; VA: vacuola. Histochemical aspects and specific stains during the ontogeny of the sporangia and receptacle scales. A. Mature receptacle scale with thickened walls (Calcofluor and epifluorescence with a DAPI-FITC-Texas filter). B. Longitudinal section of the wings from the mature receptacle scales (Toluidine blue). C-D. Mature receptacle scales with thickened cuticle (Ruthenium red). E. Sporangia and receptacle scales with polyphenols (Fast Blue B). F. Section of the sporangium annulus showing big vacuoles with polyphenols deposits (Toluidine blue). G-H. Structural and reserve carbohydrates stained Red-fuchsia alongside proteins in dark blue (PAS-Amidoblack).
AER: wings of the receptacle scales; AN: sporangium annulus; EP: sporangia; ER: mature receptacle scales; EP: spores; GA: starch granules; LF: leaf blade, PF: polyphenols; PEE: stalk of the sporangia; PER: stalk of the receptacle scales; R: receptacle; VA: vacuole.
Discusión
Los características morfológicas observadas en la presente investigación, relacionadas con el hábito, complejidad foliar, número y ubicación de los soros en la fronda, soros cubiertos con parafisis o escamas receptaculares que se pierden en la madurez, morfología y estructura de las escamas receptaculares, esporangios y ornamentación de las esporas, confirman la circunscripción taxonómica de la especie en estudio como P. macrocarpa (Hennipman et al., 1990; Otto et al., 2009; Smith &Tejero-Díez, 2014; Lagoria et al., 2018).
Las esporas maduras de P. macrocarpa observadas con luz ultravioleta no presentan afinidad por los fluorocromos para la celulosa como el Calcofluor, en cambio, muestran autoflourescencia cercana al amarillo-verdoso indicando que el esporodermo de las esporas maduras está constituido principalmente por esporopolenina. De igual manera, Rincón, Rolleri, Alzate y Dorado (2014a) llegaron a conclusiones similares en Lycopodiaceae. Por otro lado, la autofluorescencia amarillo-naranja observada en los anillos de los esporangios maduros y parte de los pedicelos indican componentes de pared secundaria, principalmente lignina y posiblemente compuestos lipídicos como la suberina, como lo reportado por Ruzin (1999) para tejidos vegetales. Los pedicelos y las células de la pared de los esporangios teñidos con Calcofluor presentan una fuerte emisión en el azul sugiriendo la presencia de componentes de pared primaria, como celulosa, siendo esto congruente con observaciones previas para este tipo de fluorocromo (Ruzin, 1999; Rincón et al., 2014a; Soukup, 2014; Yeung et al., 2015).
Bower (1963), Gifford y Foster (1989) y Kumar (2001) señalan que el desarrollo de los soros en los helechos es de tres tipos: simple, gradual basípeto y mixto o mezclado. En P. macrocarpa se evidencia el tipo de desarrollo mixto, debido a que en los soros se observaron esporangios en diferentes etapas del desarrollo. Según Schölch (2003) los soros observados en Polypodiaceae son superficiales (abaxiales) sin vascularización, lo cual discrepa con los hallazgos en la presente investigación, ya que en P. macrocarpa se observan haces vasculares en el tejido del receptáculo. Recientemente, Triana-Moreno (2012) indicó que los soros de Pecluma eurybasis var. villosa (C. Chr.) M.G. Price también presentan desarrollo mixto. El valor adaptativo de este tipo del desarrollo de los soros al parecer está relacionado con la producción de esporas en diferentes momentos ecológicos maximizando de esta manera la supervivencia de los gametófitos ante posibles adversidades climáticas. Sin embargo, estos aspectos del desarrollo de la biología reproductiva de los helechos no han sido estudiados a profundidad y se hace necesario llevar a cabo investigaciones ecológicas y reproductivas detalladas que soporten esta hipótesis o que direccionen las observaciones hacia el posible valor adaptativo de los tipos de desarrollo de los esporangios en el soro (Gifford & Foster, 1989; Qiu et al., 1995; Kumar, 2001).
Las características morfocitológicas de las células epidérmicas del receptáculo de P. macrocarpa son similares a las descritas para otras especies de helechos (Wilson, 1958; Gifford & Foster, 1989; Qiu et al., 1995; Kumar, 2001; González et al., 2010; Triana-Moreno, 2012). Sin embargo, Parkinson (1995) describe las células precursoras de los esporangios en Schizaea pectinata (L.) Sw. como protuberantes sobre la lámina foliar en desarrollo, estas son de contorno globoso y se diferencian de las demás células adyacentes. Estas características difieren de las observadas en las células epidérmicas receptaculares presentes en P. macrocarpa siendo estas alargadas y no sobresalen del receptáculo. No obstante, estas diferencias, los procesos posteriores de división mitótica que experimentan estas células son similares a lo observado en helechos leptosporangiados.
Los estudios comparativos sobre el desarrollo de los soros han mostrado que las etapas iniciales del desarrollo de los esporangios y las escamas receptaculares son similares y posteriormente divergen para la diferenciación de estas dos estructuras (Peterson & Kott, 1974). Estos resultados muestran similitud con las observaciones hechas sobre el origen y desarrollo de los esporangios y las escamas receptaculares de P. macrocarpa a partir de las mismas células del receptáculo. En este sentido las escamas receptaculares y los esporangios de P. macrocarpa se podrían considerar estructuras homólogas.
Las escamas peltadas multicelulares vegetativas están presentes en algunos grupos de helechos representantes de la familia Davillaceae, Lomariopsidaceae, Dryopteridaceae, Oleandraceae, Polypodiaceae, Cyatheaceae y Saccolomataceae (Tsutsumi & Kato, 2008). No obstante, las escamas receptaculares peltadas que cubren a los esporangios solo se han registrado en algunos géneros de Polypodiaceae (Hennipman et al., 1990, Smith &Tejero-Díez, 2014; Almeida, Salino, Dubuisson, & Hennequin, 2017) lo que se puede constatar en este trabajo para P. macrocarpa.
Tsutsumi y Kato (2008) describen detalladamente la secuencia ontogenética de la formación de las escamas peltadas multicelulares vegetativas de Nephrolepis cordifolia (L.) C. Presl., Leucostegia immersa Wall. ex C. Presl, y Davallia trichomanoides Blume. Estos autores indican que las divisiones mitóticas iniciales de las células epidérmicas receptaculares que dan origen a los tricomas o escamas multicelulares peltados se dan en el mismo plano periclinal, con lo cual se forma una estructura a manera de tricoma pluricelular simple. Posteriormente, una célula de contorno deltoide localizada en el primer tercio basal de esta estructura se divide en un plano oblicuo para formar dos células que contribuirán con la formación del escudo del tricoma. Esta secuencia del desarrollo de las escamas peltadas de estructuras vegetativas es similar a la secuencia ontogenética descrita en la presente investigación para las escamas que se diferencian en los receptáculos de P. macrocarpa. Estos hallazgos indican que el patrón de desarrollo que siguen las células epidérmicas que darán origen a los tricomas pluricelulares peltados en los órganos vegetativos en algunas especies de helechos, podría ser los mismos que presentan las células epidérmicas receptaculares. Este patrón del desarrollo al parecer determina que las células receptaculares epidémicas se diferencien en estructuras vegetativas o en estructuras reproductivas capaces de experimentar meiosis y formar esporas (Wagner, 1964). Así mismo, se confirma que las etapas iniciales de la ontogenia de las escamas multicelulares peltadas presentes en algunas especies de Bromeliaceae son notablemente similares a las observadas en P. macrcarpa (Papini, Tani, Falco, & Brighigna, 2010).
Las divisiones anticlinales y periclinales que determinan la diferenciación de los primordios de los esporangios de P. macrocarpa, así como, las características citológicas de los esporocitos premeióticos y la forma como éstos se dividen para originar al tapete celular son muy similares a las descripciones previas hechas para helechos leptosporangiados, y al parecer, son caracteres conservados en este grupo (Wilson, 1958; Peterson & Kott, 1974; Bowen & Williams, 1977; Sheffield & Bell, 1979; Qiu et al., 1995; Passarelli et al., 2010). Adicionalmente, las características citológicas de los esporocitos premeióticos de P. macrocarpa son notablemente similares a las observadas en algunas especies de Equisetum L. y en Lycopodiaceae (Rincón et al., 2011, 2013, 2014a, Rincón, Rolleri, Passarelli, Espinosa, & Torres, 2014b).
En helechos y en plantas con semilla se han identificados dos tipos principales de tapete: el celular y el plasmodial, esta estructura aporta sustancias de reserva y estructurales, contribuyendo a la síntesis de precursores de la pared de las esporas y granos de polen (Lugardon, 1990; Cresti, Blackmore, & van Went, 1992; Pacini & Franchi, 1993; Furness & Rudall, 2001; Furness, 2008; González et al., 2010; Rincón et al., 2013; Verstraete, Moon, Smets, & Huysmans, 2014). La presencia de orbículas en las cámaras plasmodiales que rodean a cada espora en maduración y la forma en que el tapete plasmodial se degenera con el desarrollo de las esporas de P. macrocarpa sugiere una función similar. Estas mismas características y funciones del tapete han sido propuestas para otras especies de pteridófitos sensu lato (Uehara & Kurita 1991; Rincón et al., 2013, 2014a, 2014b). La presencia de orbículas en especies que forman tapete plasmodial no representa una característica ampliamente distribuida, por el contrario, ésta ha sido relacionado con especies que forman tapete celular (Rincón et al., 2014a, 2014b; Verstraete et al., 2014). Se ha señalado que los pteridófitos sensu lato que forman tapete plasmodial en etapas tempranas de la ontogenia del esporangio el tapete puede estar formado de varias capas o ser biestratificado, como el observado en P. macrocarpa, en donde la capa interna del tapete adyacente a los esporocitos premeióticos pierde la integridad histológica dando origen a un plasmodio o tapete plasmodial. Este tapete invade la cavidad de los esporangios rodeando a los esporocitos y separándolos entre si y probablemente acelerando los procesos de desarrollo y diferenciación celular como lo han descrito otros autores (Wilson, 1958; Sheffield & Bell, 1979; Parkinson, 1987; Uehara & Kurita, 1989; Lugardon, 1990; Parkinson, 1995; Parkinson & Pacini, 1995; Rincón et al., 2011, 2013). No obstante, Triana-Moreno (2012) propone la presencia de una sola capa de tapete en Pecluma eurybasis var. villosa como un carácter potencialmente informativo en las relaciones intergenéricas del grupo. En relación a esto, y teniendo en cuenta la importancia que la autora da a esta estructura, se sugiere ampliar las investigaciones en esta especie para confirmar esta aseveración. De acuerdo con el análisis realizado a las imágenes que ilustran los resultados de esta autora, se observa material granular bastante regular con estructuras semejantes a núcleos dentro de la cavidad del esporangio que recuerda con las características del citoplasma del tapete plasmodial. Lo cual sugiere que el tapete en las primeras etapas del desarrollo fue biestratificado y que el estrato interno dio origen al tapete plasmodial, Furness, Rudall y Sampson (2002) indicaron que la meiosis simultánea en plantas se caracteriza porque la placa celular solo se deposita al final de la meiosis II y determina la formación de tétradas de esporas en disposición tetraedral, y que por lo general se caracterizan por formar esporas triletes. En la meiosis sucesiva, la placa celular se deposita desde la meiosis I determinando así, la formación de tétradas en disposición tetragonal, decusadas, lineal o en forma de “T” y la formación de esporas monoletes. Los hallazgos para P. macrocarpa no coinciden con estas observaciones, ya que se pudo constatar que la meiosis es de tipo simultánea, pero con la formación de tétradas en disposición decusada o tetragonal y la formación de esporas monoletes. Se verificó además en esta investigación que para P. macrocarpa, la banda de orgánulos cumple una función importante indicando el lugar exacto del depósito de la placa celular y por consiguiente en la forma como se organizan las esporas al interior de la tétrada. Estas observaciones están en concordancia con los hallazgos hechos por otros autores (Sheffield & Bell, 1979; Sheffield et al., 1983; Brown & Lemmon, 2001a, 2001b; Rincón et al., 2013).
En esta investigación se demostró la presencia de cámaras plasmodiales rodeando a los esporocitos desde el inicio de la meiosis y se mantienen alrededor de las esporas en formación hasta su madurez. Los movimientos del tapete y la formación de las cámaras plasmodiales también son similares a lo descrito para Psilotum nudum (L) P. Beav. (Psilotaceae) por Parkinson (1987), en Equisetum arvense L. (Uehara & Kurita, 1989) y Equisetum flluviatile L. (Lehmann, Neidhart, & Schlenkermann, 1984). Rincón et al. (2013) descartan la posibilidad de que las cámaras plasmodiales sean resultado del proceso de fijación de las muestras y señalan la presencia de cuerpos multivesiculares en estas cámaras que podrían estar relacionados con el tránsito de materiales desde el plasmodio hasta las esporas en maduración. En las observaciones de P. macrocarpa concordamos con estas apreciaciones relacionadas con el origen y función de esta estructura.
La síntesis del esporodermo en licófitos y monilófitos homosporados es un proceso conservado que implica primero, la formación del exosporio, luego el endosporio y al final se deposita el perisporio, cuando está presente (Lugardgon, 1990; Tryon & Lugardon, 1991; Uehara & Kurita, 1991; Rincón et al., 2014a, 2014b). Estas observaciones son congruentes a los descrito acá para P. macrocarpa. Así mismo, son similares a los registrados para licófitos y monilófitos heterospóricos en términos de desarrollo y composición química, aunque difieren ampliamente en estructura (Tryon & Lugardon, 1991; Uehara, Kurita, Sahashi, & Ohmoto, 1991; Morbelli, 1995; Wellman, 2004). El patrón de ornamentación verrugosa y la presencia de orbículas asociadas al esporodermo y presentes en la cavidad del esporangio de P. macrocarpa es coincidente con observaciones previas (Tryon & Tryon, 1982; Tryon & Lugardon, 1991; Giudice, Morbelli, Piñeiro, Copello, & Erra, 2004; Morbelli & Giudice, 2010).
Son pocos los estudios anatómicos que relacionen a las escamas pluricelulares peltadas presentes en las frondas y rizomas de algunas especies de helechos, con la función de protección e hidratación (Tsutsumi & Kato, 2008; Tejero-Díez, 2009; Hietz, 2010; Lagoria et al., 2018). No se cuenta con información relevante sobre la función de las escamas receptaculares presentes en algunas Polypodiaceae. En este orden de ideas, los hallazgos en esta investigación y que están relacionados con los patrones de fluorescencia obtenidos con el uso de Calcofluor y la tinción con rojo de Rutenio en los escudos de las escamas receptaculares inmaduras de P. macrocarpa, se relacionan directamente con la abundancia de materiales celulósicos y de pared primaria, principalmente, pectinas. Esta composición química indicaría la capacidad de hidratación de estas estructuras. John y Hasenstein (2017) utilizaron estas dos mismas técnicas de tinción y observaron un comportamiento similar, pero en las escamas multicelulares peltadas maduras presentes en las frondas de Pleopeltis polypodioides (L.) E. G. Andrews y Windham.
A medida que las escamas receptaculares de P. marcocarpa maduran, se aprecia que las paredes celulares de los escudos desarrollan engrosamientos no lignificados, los cuales excluyen el Calcofluor y emiten patrones de autofluorescencia que reflejarían su naturaleza cuticular. Estos hallazgos, no concuerda con las observaciones hechas por John y Hasenstein (2017) de las escamas multicelulares peltadas maduras de P. polypodioides, que siempre presentaron afinidad por el Calcofluor en cualquier momento del desarrollo.
En las secciones obtenidas de escamas receptaculares maduras de P. marcocarpa se observó una fuerte reacción con el azul de Toluidina indicando componentes de pared primaria y con el rojo de Rutenio que reacciona con las pectinas. Estos resultados nos permiten deducir que los escudos de las escamas receptaculares maduras además de la cutícula, presentan componentes de pared primaria que se pueden detectar debido al degaste de la cutícula producido por los solventes utilizados en el procesamiento de las muestras.
De lo anterior se podría hipotetizar que la composición química diferencial de las paredes celulares durante el desarrollo de las escamas receptaculares de P. macrocarpa estarían reflejando las necesidades fisiológicas de la planta para mantener hidratados a los esporangios inmaduros durante las primeras etapas del desarrollo. Una vez los esporangios alcanzan la madurez, las escamas receptaculares se desprenden favoreciendo la desecación de los anillos de los esporangios y la liberación de las esporas. No obstante, para apoyar esta idea se necesita realizar estudios adicionales histoquímicos, morfológicos y ultraestructurales de las escamas receptaculares en P. macrocarpa.
Los polifenoles son metabolitos secundarios abundantes en los esporangios y las escamas receptaculares en cualquier momento del desarrollo de P. macrocarpa. Estos compuestos son abundantes en las células y se almacenan principalmente en las vacuolas desplazando al citoplasma y núcleo a una posición parietal. Esta situación también la evidenciaron Peterson y Kott (1974) en su estudio sobre desarrollo de los soros y paráfisis de Polypodium virginiatum L. La presencia de estos metabolitos secundarios podría estar relacionada con la regulación de varios procesos metabólicos, con la respuesta de la planta para evitar la herbivoría y como protección contra la radiación ultravioleta, tal y como se ha descrito ampliamente para varios grupos de plantas (Kulbat, 2016; Lattanzio, Lattanzio, & Cardinali, 2006; Oszmiański, Kolniak-Ostek, & Biernat, 2015).
En P. macrocarpa, la abundancia de almidones en las células de los receptáculos durante etapas tempranas del desarrollo de los soros sugiere su utilización como fuente de energía a corto plazo y en etapas de gran actividad metabólica. Además, indica la importancia de este tejido parenquimatoso que nutre a los esporangios en maduración. No obstante, esta forma de utilización de los almidones no es exclusiva de helechos, sino de la mayoría de las plantas (Taiz, Zeiger, Møller, & Murphy 2014). Así mismo, la presencia de este compuesto en las células en los primordios de los esporangios indica la alta actividad metabólica de estas células en relación con su activo crecimiento y diferenciación.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

