











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkLos bosques andinos son sitios reconocidos como hotspots de biodiversidad por su alto grado de endemismo y alta riqueza de especies (Orme et al., 2005). Sin embargo, estos han sido fuertemente sometidos a presiones antrópicas como la deforestación, la ampliación de la frontera agrícola, el desarrollo urbanístico y la extracción o saqueo de madera, lo que ha conllevado a una pérdida del 73 % de su cobertura original desde 1980 hasta el 2006 (Cavelier & Etter, 1993; Etter, Mcalpine, Wilson, Phinn, & Possingham, 2006). Generalmente estos cambios en la cobertura arbórea, van acompañados de un proceso de fragmentación, cuya principal consecuencia es la formación de parches de bosque de diferentes tamaños aislados entre si, dando como resultado una alteración de la estructura, distribución y funcionamiento de las comunidades vegetales, además de una reducción de los tamaños poblaciones de las plantas nativas (Saunders, Hobbs, & Margules, 1991; Fahrig, 2003; Etter et al., 2006).
Esta fragmentación del bosque, puede afectar de manera particular poblaciones de especies endémicas con distribuciones restringidas y sistemas reproductivos complejos. Este es el caso del roble negro (Colombobalanus excelsa (Lozano, Hern. Cam. & Henao) Nixon & Crepet)), especie endémica de los bosques andinos colombianos que presenta un sistema sexual monoico, con inflorescencias unisexuales, baja tasa de germinación y bajo reclutamiento de nuevos individuos (Cárdenas & Salinas, 2006; Palacio-Mejía & Fernández-M,
2006; Parra-Aldana, Diez-Gómez, & MorenoHurtado, 2011). Su distribución ha quedado restringida a solo cuatro localidades, situadas entre 1 400 y 2 200 m.s.n.m en las tres cordilleras andinas. Particularmente en estas altitudes, los cambios en la cobertura original del bosque se han dado principalmente para el establecimiento de café y otros cultivos como actividad económica, y en los últimos años la pérdida de bosque en estos sitios, se ha relacionado con la ganadería extensiva y el crecimiento urbano (Etter et al., 2006; Forero, 2010).
Por lo tanto, el objetivo de este estudio se centró en evaluar la estructura poblacional y la distribución espacial de poblaciones de roble negro en cuatro fragmentos de bosque situados en la Serranía de Peñas Blancas-Huila, distribución más sur de la especie, con la finalidad de aportar conocimiento de su dinámica ecológica. Actualmente el roble negro está catalogado como vulnerable a la extinción en estado silvestre, debido a la fragmentación de bosque y al saqueo selectivo de su madera (Cárdenas & Salinas, 2006; Ministerio de Ambiente, Vivienda y Desarrollo Territorial, 2010). Según Devia y Arenas (1995) y Calderón (2001), la vulnerabilidad de esta especie está asociada particularmente a que es un árbol emergente del bosque, con características que lo hacen propicio para ser utilizado como materia prima por los aserradores locales, ya que puede alcanzar una altura entre 20 y 40 m, con copa globosa y fuste recto, de más de un metro de diámetro a la altura del pecho y una madera de alta dureza (Parra-Aldana et al., 2011). Es por esto que se espera encontrar poblaciones aisladas, en fase de recuperación, además de distribuciones no aleatorias de los individuos en cada fragmento de bosque, ya que en la zona de estudio se presenta tala selectiva y un fuerte proceso de fragmentación de bosque.
Materiales y métodos
Área de estudio
Los cuatro fragmentos de bosque estudiados La Palma, La Marimba, Alto Santa Bárbara y Alto Bellavista, se ubican en la parte más alta de la Serranía de Peñas Blancas (i.e., alturas comprendidas entre 1 630 y 1 900 m), en el extremo suroriental del departamento del Huila (Apéndice digital 1), área más sur de distribución de la especie. Esta zona corresponde al bosque sub-andino (Cleef, Rangel, Van der Hammen, & Jaramillo, 1984), comprendendiendo zonas de montaña con pendientes marcadas. La temperatura promedio de la región varía entre 16 y 20°C, la precipitación presenta un pico máximo de lluvias en el mes de julio de 235 mm y uno mínimo en el mes de enero de 63 mm (Eslava, López, & Olaya, 1986).
Muestreo
Estructura poblacional: En cada fragmento de bosque se delimitaron cuatro parcelas de 50 x 20 m (1 000 m2, Urrego & González 2000; Vallejo, Londoño-V., López-C., Galeano, Álvarez-D., & Devia, 2005) durante el año 2006, con un área total medida de 4 000 m2 (0.4 ha). En cada una, se midieron todos los individuos de roble negro sin discriminar su diámetro a la altura del pecho (DAP). Se clasificaron como: plántulas a los individuos con una altura ≤ 1.3 m, a los cuáles no se les midió DAP, sino únicamente su altura. Juveniles a los individuos con un DAP ≤ 9.9 cm, adultos a los individuos con DAP ≥ 10 cm (Caldato, Vera, & Mac, 2002). Dentro de cada fragmento de bosque, las parcelas fueron ubicadas aleatoriamente con la finalidad de obtener la mayor variabilidad local posible. Posteriormente, las cuatro parcelas por fragmento de bosque fueron sumadas a una unidad de análisis, con la finalidad de obtener la estructura etaria por fragmento de bosque y no por parcela, ya que el interés particular era comparar los diferentes fragmentos de bosque; quedando así un área total evaluada por fragmento de bosque de 4 000 m2 (0.4 ha).
Distribución espacial: Para determinar la distribución espacial del roble negro en cada parcela de 1 000 m2, cada individuo fue ubicado en un plano cartesiano, donde X indicaba una distancia de 0 a 20 m y Y de 0 a 50 m (Alder & Synnot, 1992; Vallejo et al., 2005). Para la distribución espacial tambien fueron sumadas las cuatro parcelas por fragmento de bosque, donde X indica una distancia de 0 a 40 m y Y una distancia de 0 a 100 m.
Análisis de datos
Estructura etaria: Por medio del programa Infostat versión 2012 (INFOSTAT, 2004), se obtuvieron modelos probabilísticos de distribución de frecuencias diamétricas de todos los individuos en cada uno de los cuatro remanentes de bosque. Para incluir en los análisis los individuos a los que no se les midió DAP (i.e., plántulas), se transformaron los datos sumando uno a cada diámetro (DAP + 1), luego se dividió el total de individuos en ocho clases diámetricas. Para la obtención de los modelos, se utilizaron frecuencias relativas. La frecuencia relativa se entiende como el cociente entre la frecuencia absoluta y el tamaño de la muestra. Para determinar el ajuste del mejor modelo teórico de distribución de frecuencias diamétricas, se realizó la prueba de bondad de ajuste Kolmorogov-Smirnov (Johnson, Kotz, & Balakrisnan, 1995). Por último, se comparó la densidad de individuos por hectárea y el DAP promedio de cada fragmento por medio de la prueba χ 2 (Zar, 1999). La proporción de individuos por clase diamétrica se comparó por medio de una prueba Wilcoxon no paramétrica (Zar, 1999).
Distribución espacial: Se aplicó la metodología de análisis espacial por índices de distancia, SADIE (Spatial analysis by distance Indices) por sus siglas en inglés (Perry, 1995). Con esta metodología cada individuo recibió una posición espacial a través de sus coordenadas cartesianas (X, Y) y estos se analizaron a través del indice de agregracion Ia (Perry, 1995). Donde, valores de Ia > 1 indican distribución agregada, Ia < 1 indican distribución uniforme, y Ia = 1 indican distribución aleatoria (Perry, Bell, Smith, & Woiwoid, 1996; Perry, 1998). Estos análisis fueron desarrollados con el programa SADIEShell versión 2 (Perry et al., 1996; Conrad, Perry, Woiwoid, & Alexander, 2006).
Resultados
Estructura poblacional
En total fueron censados 1 228 individuos en los cuatro fragmentos de bosque en 1.6 ha (0.4 has por fragmento de bosque). Encontrándose 105 individuos en Alto Bellavista, 399 en la Palma, 505 en Alto Santa Bárbara y 218 en La Marimba. Al llevar esta densidad de individuos por hectárea, la densidad varío entre 262.5 ha en Alto Bellavista, 997.5 ha en la Palma, 1 265 ha en Alto Santa Bárbara y 545 ha en La Marimba, encontrándose diferencias significativas entre los fragmentos de bosque (χ2 = 788.19, g.l.= 3 p < 0.001). Asimismo el DAP promedio por fragmento varió entre 15.22 y 39.27, lo que también evidencia diferencias significativas entre fragmentos de bosque (χ2= 15.35, g.l.=3 p < 0,005; Cuadro 1).
Cuadro 1 Individuos totales medidos en cada fragmento de bosque. Table 1 Total individuals measured in each forest fragment
| Fragmento de Bosque | Ind. totales | Ind. Ha | DAP prom. |
|---|---|---|---|
| Alto Bellavista | 105 | 262.5 | 39.27 |
| La Palma | 399 | 997.5 | 15.22 |
| Alto Santa Bárbara | 506 | 1 265 | 17.14 |
| La Marimba | 218 | 545 | 22.10 |
Ha: hectárea, DAP: dimátro a la altura del pecho promedio para cada fragmentos de bosque.
Cuadro 2: Densidades de los individuos de roble negro para los cuatro fragmentos de bosque. Table 2: Densities of black oak individuals for the four forest fragments
| - | Alto Bellavista | - | - | Alto Santa Bárbara | - | ||||
|---|---|---|---|---|---|---|---|---|---|
| Clases diamétricas | Frecuencia absoluta | Frecuencia relativa | Individuos por Ha | Clases diamétricas | Frecuencia absoluta | Frecuencia relativa | Individuos por Ha | ||
| 1-35.88 | 62 | 0.59 | 155.00 | 1-25.75 | 406 | 0.8 | 1 015 | ||
| 35.88-70.75 | 20 | 0.19 | 50.00 | 25.75-50.5 | 62 | 0.12 | 155 | ||
| 70.75-105.63 | 13 | 0.12 | 32.50 | 50.5-75.25 | 28 | 0.06 | 70 | ||
| 105.63-140.5 | 8 | 0.08 | 20.00 | 75.25-99.99 | 5 | 0.01 | 12.5 | ||
| 140.5-175.38 | 2 | 0.02 | 5.00 | 99.99-124.74 | 4 | 0.01 | 10 | ||
| 175.38-210.25 | 0 | 0 | 0 | 124.74-149.49 | 0 | 0 | 0 | ||
| 210.25-245.13 | 0 | 0 | 0 | 149.49-174.24 | 0 | 0 | 0 | ||
| 245.13-280 | 0 | 0 | 0 | 174.24-198.99 | 1 | 0 | 2.5 | ||
| total | 105 | 1 | 262.5 | total | 506 | 1 | 1 265 | ||
| - | La Palma | - | - | La Marimba | - | ||||
| Clases diamétricas | Frecuencia absoluta | Frecuencia relativa | Individuos por Ha | Clases diamétricas | Frecuencia absoluta | Frecuencia relativa | Individuos por Ha | ||
| 1-34.82 | 348 | 0.87 | 870 | 1-23.2 | 153 | 0.7 | 382.5 | ||
| 34.82-68.64 | 35 | 0.09 | 87.5 | 23.2-45.4 | 28 | 0.13 | 70 | ||
| 68.64-102.46 | 9 | 0.02 | 22.5 | 45.4-67.61 | 10 | 0.05 | 25 | ||
| 102.46-136.28 | 4 | 0.01 | 10 | 67.61-89.81 | 13 | 0.06 | 32.5 | ||
| 136.28-170.1 | 0 | 0 | 0 | 89.81-112.01 | 5 | 0.02 | 12.5 | ||
| 170.1-203.92 | 1 | 2.50E-03 | 2.5 | 112.01-134.21 | 4 | 0.02 | 10 | ||
| 203.92-237.74 | 0 | 0 | 0 | 134.21-156.41 | 4 | 0.02 | 10 | ||
| 237.74-271.56 | 2 | 0.01 | 5 | 156.41-178.62 | 1 | 4.60E-03 | 2.5 | ||
| Total | 399 | 1 | 997.5 | Total | 218 | 1 | 545 | ||
La mayor proporción de individuos en los cuatro fragmentos de bosque, se encuentra en las tres primeras clases diamétricas. Alto Bellavista presentó en su primer clase diamétrica 59 % del total de los individuos censados, Alto Santa Bárbara 80 %, La Palma 87 % y La Marimba 70 % respectivamente (Cuadro 2). La comparación entre fragmentos de bosque, evidencia que no existen diferencias significativas entre la proporción de individuos en cada clase diamétrica (W= 6.02, p = 0.1), independientemente de la cantidad de individuos encontrados en cada fragmento de bosque.
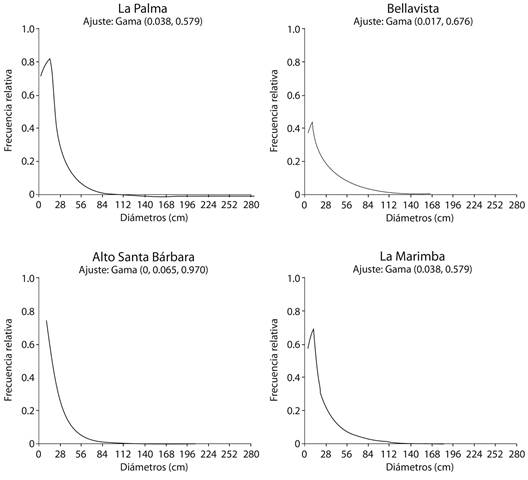
Modelos probabilísticos y estructura etaria: Para cada uno de los fragmentos de bosque se obtuvo un modelo de distribución de frecuencias diamétricas (Apéndice digital 2, Figura 1). Estos modelos tuvieron dos tipos de ajustes para los cuatro fragmentos de bosque estudiados, así el modelo probabilístico de distribución de frecuencias diamétricas con mayor ajuste fue el Gamma, para los fragmentos de bosque Bellavista, La Palma y La Marimba; y, el modelo Weibull, para el fragmento de bosque Alto Santa Bárbara (Figura 1). Los modelos Gamma y Weibull que se ajustaron a cada fragmento, sugieren que los bosques pertenecen a rodales multietáneos.
Distribución espacial del roble negro en cada remanente de bosque: Todos los fragmentos presentaron distribuciones espaciales agregadas Ia > 1 aunque con diferentes valores de significancia, así: Alto Bellavista (Ia= 1.48, p= 0.04) y La Marimba (Ia= 1.87, p= 0.003) presentaron una distribución agregada estadísticamente significativa y, La Palma (Ia= 10.97, p= 0.0002) y Alto Santa Bárbara (Ia= 10.85, p= 0.0002) presentaron una distribución agregada altamente significativa.
Discusión
Existe una gran variación en la densidad de individuos por hectárea para cada uno de los fragmentos de bosque. Esta gran variación podría ser el resultado de diferentes procesos de regeneración y establecimiento después de diferentes intensidades de tala o saqueo de madera. Por ejemplo, en el fragmento de bosque de Alto bellavista, el cual es protegido por la comunidad local y tuvo una menor intensidad de saqueo, la densidad fue de 262.5 ind/ ha, en contraste con el fragmento de bosque de Alto Santa Bárbara donde la densidad fue de 1 265 ind/ha siendo el fragmento de bosque más intervenido. Estos resultados coinciden con lo encontrado en otros estudios realizados sobre la misma especie y en bosques cercanos a los sitios de estudio. Por ejemplo Parra-Aldana et al. (2011), en poblaciones muy intervenidas encontraron una densidad de 430 ind/ha y Dávila, Alvis y Ospina (2012) encontraron una densidad de 281.7 ind/ha en poblaciones menos intervenidas que se encuentran en el Parque Nacional Natural Cueva de los Guácharos.
En general el mayor porcentaje de individuos en todos los fragmentos de bosque se encuentra en las primeras clases diamétricas (Parra-Aldana et al., 2011; Dávila et al., 2012; Moreno, 2014) y este porcentaje es proporcional a la densidad de individuos por hectárea, e.g., Alto Bellavista tiene el 59 % del total de los individuos en la primer clase diametrica y Alto Santa Bárbara tiene el 80 %. Estos resultados indican, que estos bosques en general se encuentran en una fase de recuperación después de una fuerte intervención antrópica de diferentes intensidades en cada fragmento de bosque.
Encontrar un alto porcentaje de individuos en la primera clase diamétrica, indica que se puede estar facilitando el establecimiento de plántulas en los cuatro fragmentos de bosque, y que éste establecimiento es mayor en bosques más saqueados o talados. Probablemente existe mayor establecimiento debido a que el roble negro es una especie heliófita en sus primeros estadios de desarrollo, es decir, plántulas y árboles jóvenes necesitan de luz directa para su crecimiento y desarrollo (Nixon & Crepet, 1989; Parra-Aldana et al., 2011). Por otra parte, encontrar una alta densidad de individuos en la primera clase diamétrica en los cuatro fragmentos de bosque, permite inferir que existe una reserva de individuos jóvenes que más adelante podrían reemplazar a los árboles de mayor tamaño que vayan desapareciendo (Villa, 2005), ya sea por muerte natural o por extracción selectiva. Asimismo, la alta reserva de individuos en las primeras clases diamétricas, indica que de cierta forma la explotación selectiva de bosques de roble ha sido una herramienta eficaz para promover su regeneración natural (Guariguata & Saenz, 2002; Guerrero, Paz, & Parrado, 2010).
Sin embargo, estas conclusiones deben tomarse con precaución, ya que en este trabajo no se evaluó como varía el porcentaje de establecimiento y viabilidad de semillas. Por ejemplo, Parra-Aldana et al. (2011) encontraron un bajo establecimiento y una baja viabilidad de las semillas de la especie y Moreno (2014), encontró que la baja viabilidad de las semillas estaba asociada a una alta depredación de estas en el banco de semillas. Otros trabajos también han mencionado la dificultad de regeneración de diferentes especies de robles (González, Ochoa, Ramírez, & Quintana, 1995; Pulido, 2002), estableciendo que la falta de reclutamiento puede conducir a la disminución a largo plazo de las poblaciones naturales (Sork et al., 2002).
Los modelos de distribución de frecuencias diamétricas, los cuales incluyen la cantidad de individuos por clases de tamaños o clases diamétricas dentro de una población, son buenos predictores del estado actual de poblaciones con algún grado de amenaza, como lo es el roble negro. En este trabajo, en tres fragmentos de bosque se ajustaron modelos de distribución de frecuencias diamétricas Gamma y para el fragmento de bosque Alto Santa Bárbara, que presentó mayor disturbio antrópico, se ajustó un modelo Weibull. Esta última población (i.e., Alto Santa Bárbara), presentó mayor cantidad de individuos totales (N = 506) y mayor porcentaje de individuos en las primeras clases diamétricas (80 % de individuos en la primera clase diamétrica), en relación a los otros tres bosques fragmentados, lo cuál hizo que la curva del modelo tuviera una pendiente mucho más pronunciada. En general, estos modelos representan poblaciones con distribuciones en J invertida, que indican que se trata de poblaciones disetáneas con estructuras de tamaños aparentemente balanceadas, en las cuales existe una mayor cantidad de individuos en las categorías diamétricas menores.
La distribución espacial fue agregada (Ia > 1) para cada uno de los cuatro fragmentos de bosque, siendo altamente significativa para La palma y Alto Santa Bárbara. La distribución espacial podría estar relacionada a la densidad de individuos encontrados en las primeras clases diamétricas y esta alta densidad en las primeras clases, a su vez puede estar relacionado a un alto establecimiento de plántulas en zonas de clareo después de un proceso de tala selectiva de individuos adultos de gran porte. La tala selectiva se practicó intensivamente en la zona de estudio en los años 80.
Se puede concluir que encontrar una estructura etaria que corresponde con poblaciones en J invertida y con distribuciones espaciales agregadas, que a su vez se ajustan a modelos de distribución diamétrica Gamma y Weibull; son poblaciones que se encuentran en fase de recuperación después de una fuerte intervención antrópica. Esta intervención ha afectado de manera más drástica a las poblaciones de los fragmentos La Palma y Alto Santa Bárbara y en menor medida a las poblaciones de los fragmentos Alto Bellavista y La Marimba. Sin embargo, esta intervención antrópica ha permitido que exista un establecimiento de plántulas en zonas de clareo, que de no existir probablemente la estructura etaria correspondería a poblaciones con mayor cantidad de individuos adultos, poblaciones clímax donde el establecimiento tiende a ser minimo por la estructura de edades de los individuos (Crawley, 2003).
Finalmente, una de las consecuencias más agresivas de la fragmentación es la disminución del tamaño poblacional y los procesos que estos conllevan como son la endogamia y la disminución del tamaño poblacional efectivo. Esta especie aunque presenta una gran cantidad de plántulas y juveniles que aseguran su permanencia en el tiempo, también presenta baja diversidad genética (Aguirre-Acosta, Palacio-Mejía, Barrios-Leal, & Botero-Echeverri, 2013; Palacio-Mejía & Fernández-M., 2006), lo que podría ocasionar una progresiva disminución de la capacidad reproductiva de la especie (Swaine, Lieberman, & Putz, 1987; Parra-Aldana et al., 2011) y a su vez, una extinción a largo plazo. Es por esto, que el conocimiento que se pueda generar en cuanto a la dinámica poblacional del roble negro en cada una de las localidades donde aún existe, sirve como insumo para generar planes de manejo y conservación en conjunto con las comunidades locales.

