Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkLas actividades humanas han reducido el tamaño de las poblaciones de mamíferos silvestres y sus ámbitos de distribución, a nivel global, a tal grado, que han colocado en riesgo de extinción a una cuarta parte del total de las especies de este grupo (Ceballos & Ehrlich, 2002; Baillie, Griffiths, Turvey, Loh, & Collen, 2010). En el neotrópico las especies de la familia Felidae son de las más afectadas por las actividades humanas, ya que además de la destrucción de su hábitat, están consideradas como una amenaza para el ganado, son vendidas como mascotas y sus pieles son comercializadas (Ceballos, List, Medellín, Bonacic, & Pacheco, 2010a).
En México habitan seis de las doce especies de felinos presentes en el continente americano: lince (Lynx rufus), puma (Puma concolor), jaguarundi (Puma yagouaroundi), ocelote (Leopardus pardalis), tigrillo o margay (Leopardus wiedii) y el jaguar (Panthera onca) (Ceballos et al., 2010a). Sin embargo, no se cuenta con información precisa y actualizada sobre sus patrones de distribución a nivel local y regional. Asimismo, se desconoce en gran medida el efecto que las variables ambientales tienen sobre sus patrones de distribución (Leopold, 1959; Emmons & Feer, 1997).
La vertiente del Pacífico es una región de México que en particular carece de información detallada sobre la presencia y distribución de especies de felinos, particularmente en la zona delimitada por el estado de Michoacán. Esto se debe en parte a la dificultad de acceso a esta región, caracterizada por una compleja topografía, consecuencia de la convergencia del Eje Neovolcánico, la Sierra Madre del Sur y la depresión de las cuencas del Balsas y Tepalcatepec (Garduño-Monroy, 2005). De acuerdo con Leopold (1959), las seis especies de felinos presentes en México se distribuían ampliamente en Michoacán. Sin embargo, la información disponible para sustentar esa afirmación era muy limitada y aunque en las décadas siguientes se agregaron nuevos registros de felinos en la región, la información es aún insuficiente, dispersa geográficamente y en algunos casos imprecisa (Brand, 1960; Álva-rez, Arroyo-Cabrales, & González-Escamilla, 1987; Olalde-García, 1997).
La región delimitada por el estado de Michoacán estuvo originalmente dominada por el bosque tropical caducifolio o (selva baja caducifolia), en combinación con el bosque tropical subcaducifolio (selva mediana) en las áreas más húmedas, y bosques templados de encino-pino en elevaciones superiores a los 1 500 msnm (Ceballos, García, Salazar, & Espinoza, 2010b). Sin embargo, la creciente actividad humana ha provocado que el paisaje de la región se caracterice en la actualidad por la presencia de un complejo mosaico de diferentes tipos de cobertura y usos del suelo, tales como bosques tropicales y bosques templados altamente fragmentados por la agricultura y ganadería (Bocco, Mendoza, & Masera, 2001). El creciente impacto humano sobre los hábitats naturales existentes en la región aumenta la importancia de llenar el vacío de información existente sobre los patrones actuales de distribución de las especies de felinos.
Por lo anterior, los objetivos de este estudio se centraron en: 1) recabar y analizar la información existente sobre la distribución de las especies de felinos en la Costa del Pacífico, en particular en la región delimitada por el estado de Michoacán; 2) actualizar la información sobre la distribución actual de los felinos, a partir de la integración y análisis de los registros obtenidos para este grupo mediante trabajo de campo sistemático realizado durante los últimos 10 años; y 3) analizar cómo se distribuyen las especies en relación con la existencia de distintas ecorregiones, tipos de vegetación y variaciones climáticas. Todo con el fin de avanzar en el conocimiento de la ecología y estado de conservación de los felinos en el trópico seco del Pacífico Central.
Materiales y Métodos
Área de estudio: El estado de Michoa-cán se localiza entre las coordenadas 17°55' - 20°24' N y 100°04' - 103°44' W, y abarca una superficie de 58 364 km2 (Garduño-Monroy, 2005). Debido a su heterogeneidad topográfica, altitudinal (0 hasta 3 569 msnm) y de tipos de vegetación, en el estado se han identificado las siguientes ecorregiones (Olson et al., 2001; Garduño-Monroy, 2005): Bajío, Eje Neovolcá-nico, Bajo Balsas, Sierra Madre del Sur y Costa. En la región del Bajío, predomina el matorral subtropical y extensas planicies; en el Eje Neo-volcánico, predominan los bosques templados de coníferas y de encino. En las ecorregiones del Bajo Balsas, Costa y Sierra Madre del Sur, dominan los bosques tropicales caducifolios y subcaducifolios, con una menor representación de bosques templados típicos de elevaciones superiores (Carranza-González, 2005).
Integración de registros de presencia de felinos en la región de Michoacán a partir de referencias bibliográficas y bases de datos: Se revisaron y consultaron artículos científicos, tesis de grado y bases de datos nacionales como la Red Mundial de Información sobre Biodi-versidad REMIB (http://www.conabio.gob.mx/ remib/doctos/remib_esp.html) y la de la Unidad de Informática para la Biodiversidad UNIBIO (http://unibio.unam.mx/). Adicionalmente, también se consultaron portales internacionales como el: Global Biodiversity Information Facility GBIF fhttp://www.gbif.org/) y Mammal Networked Information System MaNIS (http:// manisnet.org/). Dentro de las fuentes consultadas fueron particularmente importantes los registros de referencia provenientes de los proyectos: "Mamíferos de México", del cual los especímenes fueron depositados en colecciones de Estados Unidos y Canadá (López-Wilchis, 1998), "Biodiversidad de los Mamíferos del Estado de Michoacán" en Álvarez-Solórzano y López-Vidal (1998) y el proyecto "Computari-zación de las colecciones del museo de Zoología Alfonso L. Herrera para su incorporación a la REMIB: Fase I", en León-Paniagua (1999). Todos los registros que se obtuvieron fueron revisados, verificados y corregidos, en caso de ser necesario, para asegurarse que no hubiera errores en sus coordenadas geográficas e información de localidad de procedencia.
Registros de presencia de especies de felinos obtenidos en campo (2003-2013): A partir del 2003 se inició un programa para el registro de felinos en sitios seleccionados con base en su accesibilidad geográfica, tipo de vegetación, seguridad para el acceso y apoyo de los pobladores locales.
La recolecta de registros sobre la presencia de felinos se realizó a través de: 1) registros directos (i.e., observaciones) e indirectos (i.e., cráneos, otros restos biológicos, excretas y huellas), ambos obtenidos durante recorridos diurnos y nocturnos de transectos; 2) cámaras-trampa, y 3) la realización de entrevistas estructuradas. Los sitios fueron visitados una sola ocasión, con la excepción de algunas localidades ubicadas en la Costa y la Sierra Madre del Sur 2010 y el 2013.
Los transectos tuvieron una longitud de 1.5 a 2.5 km. El 50% de los recorridos de estos tran-sectos fueron diurnos (6:00 a 10:00hr), el 25% nocturnos (22:00-2:00hr) y el 25% restante se realizó durante el crepúsculo (17:00-20:00hr). Las huellas se identificaron utilizando guías de campo y el apoyo de asistentes locales con experiencia (Aranda, 2000). Los cráneos encontrados se depositaron en las colecciones de mamíferos del Museo de Zoología "Alfonso L. Herrera" de la Facultad de Ciencias de la Universidad Nacional Autónoma de México y en la Facultad de Biología de la Universidad Michoacana de San Nicolás de Hidalgo para que fueran catalogados.
Las cámaras-trampa se colocaron al borde de senderos, caminos y manantiales con el fin de maximizar la probabilidad de registrar la presencia de felinos (Silver et al., 2004). Este muestreo, se realizó principalmente después de la época de lluvias (noviembre) y durante la época de secas (diciembre a mayo). Se utilizaron los siguientes modelos de cámaras-trampa: Moultrie Game Spy, 4.0 megapixels (Moultrie feeders, Inc. Alabaster, Alabama) y Wildview Xtreme 4 y 5 (Wildview Co., Grand Prairie, Texas). Se consideraron como registros independientes los obtenidos para la misma especie y estación de cámara-trampa pero obtenidas con una diferencia mínima de 24 horas o cuando se registró más de un individuo en una fotografía (Botello, Monroy, Illoldi-Ran-gel, Trujillo-Bolio, & Sánchez-Cordero, 2007; Monroy-Vilchis, Zarco-González, Rodríguez- Soto, Soria-Díaz, & Urios, 2011).
Para identificar los felinos registrados por fotografías (Fig. 1), se empleó la guía para mamíferos del Neotrópico de Emmons y Feer (1997). Además, para diferenciar especies similares de felinos, como L. wiedii y L. pardalis se consultó respectivamente a Murray y Gardner (1997) y Oliveira (1998).

Las entrevistas se aplicaron a los habitantes de las localidades muestreadas y sirvieron principalmente para identificar áreas con presencia de los felinos, las cuales fueron visitadas para registrar sus coordenadas. En ocasiones las entrevistas también sirvieron para obtener evidencia, tal como la piel de felinos cazados, que sirvió para confirmar la presencia de las especies.
Todos los registros bibliográficos y los obtenidos en campo durante este trabajo, están disponibles para quien los requiera a petición expresa dirigida al primer autor de este trabajo.
Análisis de los patrones de distribución de las especies de felinos: Una vez que las ncoordenadas de todos los registros de felinos se verificaron, se incorporaron a un sistema de información geográfica (SIG) creado con el programa ArcGis versión 9.3 (ESRI, 2008). En este SIG también se incorporaron los mapas de las ecorregiones propuestas por Olson et al. (2001) y de la vegetación y uso de suelo a escala 1:250 000 (INEGI, 2013a), así como los polígonos de las áreas naturales protegidas de carácter estatal y federal (CONANP, 2013). En el caso de la cartografía de vegetación y uso del suelo se combinaron las siguientes categorías: pastizales inducidos, pastizales cultivados, agricultura de temporal y agricultura de riego, para formar una nueva categoría que se denominó: Usos Agropecuarios. Finalmente, al SIG se le incorporaron las siguientes capas de información en formato raster: modelo digital de elevación (MDE) con una resolución de 30 metros por pixel (INEGI, 2013b), y variables bioclimáticas relacionadas con la precipitación y temperatura que se descargaron del sitio web WorldClim-Global Climate Data (www. worldclim.org). En específico se incorporaron las siguientes variables bioclimáticas: BIO1 = temperatura media anual, BIO5 = temperatura máxima de mes más caluroso, BIO6 = temperatura mínima de mes más frío, BIO12 = precipitación anual, BIO13 = precipitación de mes más húmedo y BIO14 = precipitación de mes más seco. Estas variables han sido asociadas con la distribución de los felinos en otras regiones (Jiménez-Maldonado, 2007; Bárcenas & Medellín, 2010; Cuervo-Robayo & Monroy-Vilchis, 2012).
Para evaluar si existía una distribución diferencial de las especies de felinos en función de las características del ambiente, se realizó un análisis de escalamiento multidimensional no métrico (NMDS). Se usó el comando veg-dist de la biblioteca Vegan del programa R (R Core Team, 2013) para calcular las distancias euclidianas entre los registros de las especies en un espacio multivariado definido por las variables climáticas antes mencionadas y la elevación. Posteriormente, usamos el comando MetaMDS para realizar el análisis de NMDS.
Este comando transforma automáticamente los datos buscando mejorar la calidad de la ordenación y busca de manera iterativa la solución del análisis con menor estrés (i.e., donde la ordenación representa de manera más fiel las distancias originales en el espacio multivariado) (Borcard, Gillet, & Legendre, 2011).
Por último, se realizó un análisis de conglomerados para determinar si distintas áreas dentro de la región de estudio, representadas a partir de distintas combinaciones de ecorre-giones y tipos de vegetación, se agrupaban en función de los patrones de presencia y tasas de captura de las especies de felinos. Se asignaron las siguientes claves de identificación a las ecorregiones (E1 Bajío, E2 Eje Neovolcánico, E3 Bajo Balsas, E4 Sierra Madre del Sur y E5 Costa) y tipos de vegetación (V1 bosque tropical caducifolio, V2 bosque tropical sub-caducifolio, V3 bosque de pino, V4 bosque de pino-encino, V5 bosque de encino, V6 área agropecuaria). Posteriormente, se calcularon las tasas de captura (no. de registros/100 días cámara-trampa) de cada especie para cada combinación ecorregión-tipo de vegetación. Con base en estos datos se calculó la distancia euclidiana entre las distintas áreas, usando el comando vegdist de la biblioteca Vegan del programa R (R Core Team, 2013). Se transformaron los datos (normalización) con la función decostand de Vegan. Se probaron distintas técnicas de aglomeramiento jerárquico para obtener un dendrograma incluyendo: ligamiento sencillo, ligamiento completo, UPGMA y Ward (para consultar detalles de estas técnicas ver Borcard et al., 2011). Para evaluar cuál de los dendrogramas resultantes era más efectivo para representar las distancias euclideanas entre las distintas áreas, se realizaron correlaciones cofenéticas (Borcard et al., 2011). Para determinar el número óptimo de conglomerados en el dendrograma se utilizó el comando silhouette de R (Borcard et al., 2011). Para este análisis sólo utilizamos los registros de cámaras-trampa por ser los más directamente comparables entre especies.
Resultados
Registros bibliográficos: Se reunieron 29 registros de los felinos (Fig. 2): P. onca (n = 13), L. rufus (n = 4), P. yagouaroundi (n = 4), L. wiedii (n = 4), P. concolor (n = 2) y L. pardalis (n = 2). Sin embargo, la mayoría de los registros carecían de una adecuada información que permitiera establecer de manera precisa sus coordenadas de origen.
Las fechas de los registros de origen bibliográfico, abarcaron desde el lince recolectado por el "Smithsonian Institution" en el municipio de Pátzcuaro en 1892 (López-Wilchis, 1998), hasta la fotografía del tigrillo obtenida mediante una cámara-trampa en el municipio de Uruapan en 2005 (Chávez-León & Zaragoza-Ribera, 2009).

Fig. 2: Geographic distribution of felid species records in the study area. Black dots correspond to records obtained from literature and data bases whereas white dots were obtained during field work conducted for this study. The gray scale represents the five ecoregions occurring in the state, from north to south; Bajio, Eje Neovolcánico, Bajo Balsas, Sierra Madre del Sur and Costa.
Fueron muy poco los casos en los que se contó con especímenes recolectados que estuvieran depositados en colecciones científicas. En ese sentido, destacan tres registros de L. rufus (López-Wilchis, 1998), uno de P. yagouaroundi (Álvarez-Solórzano & López-Vidal, 1998), y uno de L. wiedii (León-Paniagua, 1999).
La mayoría de los 29 registros bibliográficos correspondieron a reportes circunstanciales. Por ejemplo, se encontraron 13 registros bibliográficos de P. onca, que carecían de una precisa georreferenciación de respaldo, una condición que dificultó la verificación o confirmación de los registros (Núñez, 2007; 2012). Entre los registros destacan por su ubicación los reportados por Brand (1960), con coordenadas correspondientes a la cabecera municipal de Coalcomán, así como el jaguar identificado en 1987, por medio de una piel encontrada en una curtiembre del municipio de Arteaga (Núñez, 2012).
Del total de estos registros, diez se presentaron en zonas urbanas, nueve en áreas de usos agropecuarios, tres en bosques de pino, tres en bosque de pino-encino, tres en bosque tropical subcaducifolio y finalmente uno en bosque tropical caducifolio. Con respecto a las ecorregio-nes, no se encontraron registros bibliográficos para el Bajío (Fig. 2). En contraste, en la eco-rregión Costa, se concentraron once registros. El 55% de los registros se encontraron a una altura por debajo de 1 000 msnm.
Registros obtenidos en campo: Se acumuló un esfuerzo de muestreo de 1 107.5 km de transectos y 8 699 días/cámara-trampa, que produjeron un total de 672 registros de felinos en 302 localidades distintas (Fig. 2). Se obtuvieron registros con todos los métodos (Cuadro 1), aunque el más eficiente fue el uso de cámaras-trampa, que generó el 74.3% de los registros, seguido por la identificación de huellas (11.3%) y las entrevistas (9.5%). La especie con el mayor número de registros totales fue L. pardalis (51.1%), seguida de P. concolor (29.8%). En contraste, L. rufus fue la especie con el menor número de registros (0.4%). Con respecto a los registros obtenidos mediante cámaras-trampa, el ocelote presentó la tasa de captura más alta con 29.8 registros por 100 días/cámara-trampa, seguido del puma con 14.9, tigrillo con 10.7, jaguarundi con 9.8, jaguar con 3.6 y finalmente el lince con 3.3.
Se recolectaron cinco cráneos que se depositaron en colecciones científicas (Cuadro 1). Estos cráneos correspondieron a las siguientes especies: L. pardalis (MZFC 8888), P concolor (MZFC 8897), P. yagouaroundi (MZFC 8890 y UMSNH 3413) y P. onca (MZFC 11063).
Patrones de distribución de las especies de felinos: La escasez de registros de felinos obtenidos a partir de la revisión de bibliografía y de bases de datos limitó la posibilidad de hacer una descripción detallada de la distribución de las especies de felinos. Resalta, sin embargo, el que 65% de estos registros, corresponden a sitios que actualmente están sujetos a usos agropecuarios o son zonas urbanas.

Cuando se analizaron en conjunto todos los registros, destacaron el puma, ocelote y tigrillo como las especies con una distribución más amplia, que incluyó las ecorregiones de Bajo Balsas, la Sierra Madre del Sur y Costa. En estas regiones se registró en menor medida al jaguar y jaguarundi. Sin embargo, fue la ecorregión de la Sierra Madre del Sur donde se encontró el mayor número de registros, (n = 271). En contraste, la ecorregión del Bajío es donde se presentó el menor número de registros (n = 3), que sólo incluyeron a L. rufus (Fig. 2).
En general, los registros de felinos se distribuyeron en un gradiente altitudinal entre los 3 y 2 746 msnm. Sin embargo, un 89.5% de los registros se localizó por debajo de los 1 000 msnm. L. rufus fue la especie cuyos registros tuvieron una distribución altitudinal más restringida ya que sólo se encontraron entre los 1 700 y 2 700 msnm. En contraste, P. concolor y L. wiedii fueron las especies que se registraron en el intervalo altitudinal más amplio (3 a 1 758 msnm).
El clima de la región donde se localizaron los 701 especímenes de felinos, presentó una precipitación durante el mes más seco de menos de 10 mm, y el 82% (n = 579) de los registros, correspondieron a zonas con escasa precipitación, pero con una precipitación en el mes más húmedo entre 201 y 300 mm. El 56.0% (n = 392) de los registros se presentaron en áreas con una temperatura promedio anual entre 21 y 25°C. El 71% (n = 496) de los registros se localizaron en áreas donde la temperatura del mes más cálido fluctuó entre los 31 y 35 °C. Por otra parte, el 69% (n = 483) de los registros, se presentó en áreas donde la temperatura más fría varió entre 16 y 20°C. En la mayoría de especies, a excepción del lince y el jaguar, los registros se presentaron en localidades donde la temperatura máxima fue superior a los 39 °C.
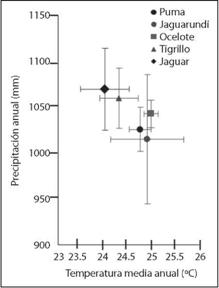
En general, los felinos se registraron en sitios que presentaron una temperatura media anual de 24 °C y una precipitación anual de 1 040 mm. La única excepción fue L. rufus, que se registró a una temperatura promedio de 15.8 ± 1.3 °C y una precipitación media anual de 941 ± 171 mm (Apéndice 1).
Los resultados del análisis de NMDS hicieron evidente que la mayoría de las especies de felinos presentaron una afinidad con respecto a las condiciones ambientales de las localidades en las que hubo evidencia de su presencia (Fig. 3). La única excepción fue el lince, ya que las localidades donde se presentó se separaron claramente en el NMDS en particular por estar ubicadas a una mayor altitud y una precipitación del mes más seco, superior a los 6 mm (Fig. 3).
Al analizar la tasa de registro de los felinos en las cámaras-trampa, encontramos que en la ecorregión del Bajío solo se registró al lince con una tasa de captura de 3.3 registros por 100 días/cámara-trampa. Mientras que en el bosque tropical caducifolio de la Costa y en los bosques de pino-encino de la Sierra Madre del Sur se registraron la mayor cantidad de especies de felinos con cinco y cuatro y las mayores tasas de captura de felinos con 9 y 8.5 registros por 100 días/cámara-trampa, respectivamente.
De acuerdo con el dendrograma resultante del análisis de conglomerados de las combinaciones de ecorregiones y tipos de vegetación se logró distinguir cinco grupos (Fig. 4A). El grupo uno, incluyó a cinco áreas dentro de las ecorregiones del Eje Neovolcánico, Bajo Balsas y Costa, donde se registraron a cinco especies de felinos (sólo el lince estuvo ausente), con una tasa de captura de 15.3 registros por 100 días/cámara-trampa para los felinos registrados en bosques tropicales caducifo-lios, 3.8 registros por 100 días/cámara-trampa para los felinos registrados en bosque tropical subcaducifolio, y 13.7 registros por 100 días/ cámara-trampa para los felinos registrados en bosques de pino-encino templada (Fig. 4A, Fig. 4B). El grupo dos, incluyó a registros de jaguarundi, ocelote y tigrillo en bosques tropicales caducifolios de la Sierra Madre del Sur con 7.6 registros por 100 días/cámara-trampa, terrenos agropecuarios en la Costa con 0.8 registros por 100 días/cámara-trampa y bosques de pino en el Eje Neovolcánico con 1.7 registros por 100 días/cámara-trampa (Fig. 4A, Fig. 4B). El grupo tres incluyeron los registros de puma, ocelote, tigrillo y jaguar para cuatro áreas: bosque tropical caducifolio del Eje Neovolcánico con 6.2 registros por 100 días/cámara-trampa, usos agropecuarios del Bajo Balsas con 3.1 registros por 100 días/cámara-trampa, bosques de pino de la Sierra madre del Sur con 1.5 registros por 100 días/cámara-trampa y bosque tropical subcaducifolio de la Costa con 2.6 registros por 100 días/cámara-trampa (Fig. 4A, Fig. 4B). El grupo cuatro incluyó únicamente a los bosques de encino de la Sierra Madre del Sur, donde se presentaron tasa de captura de 2.5, 7.5 y 2.5 registros por 100 días/cámara-trampa para puma, jaguarundi y ocelote, respectivamente (Fig. 4A, Fig. 4B). Por último, el quinto grupo incluyó a los dos registros de cámara-trampa de lince, los cuales se obtuvieron en los bosques tropicales caducifolios del Bajío, con una tasa de captura de 3.3 registros por 100 días/cámara-trampa (Fig. 4A, Fig. 4B).
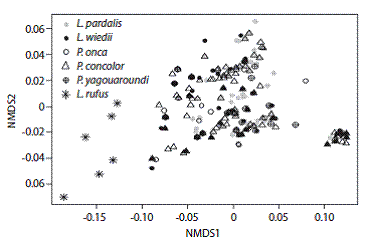
Fig. 3 Analysis of non-metric multidimensional scaling depicting the distribution of felid presence records in relation to a set of seven environmental variables. There is a general overlap in the distribution of the species with the exception of Lynx rufus which shows the most distinctive pattern.
Sólo 44 de los 701 registros correspondieron a sitios dentro de alguna de las 32 áreas naturales protegidas de carácter estatal y 13 de carácter federal existentes en la región. Prácticamente, la totalidad de estos registros se presentaron dentro de la Reserva de la Biosfera de Zicuirán Infiernillo (RBZI), y correspondieron a: P. onca (n = 1), P. concolor (n = 22), P. yagouaroundi (n = 3), L. pardalis (n = 15) y L, wiedii (n = 2). L. wiedii también se registró en el Parque Nacional Barranca del Cupatitzio. El lince fue la única especie para la que no se obtuvieron registros en alguna área protegida.
Discusión
El trabajo de campo desarrollado en los últimos 10 años permitió pasar de 29 a 672 registros de felinos en la región de estudio, lo que significa un incremento de más del 2 000% en la información disponible. A partir de esta información es posible tener una visión más completa de los patrones de distribución de las especies de felinos en esta importante porción de la costa del Pacífico mexicano.
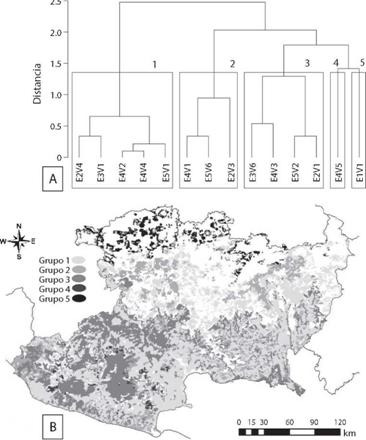
Fig. 4: A) Cluster analysis showing that it is possible to distinguish five groups of areas having different combinations of vegetation types and eco-regions based on the camera-trap records of the different felid species (key for different combinations of eco-regions and vegetation types is explained in the methods section). B) Geographic location of the five clusters.
Los resultados muestran que las cinco especies de felinos de afinidad tropical presentan una distribución relativamente extensa que abarca un amplio intervalo altitudinal (desde el nivel del mar hasta los 1 760 msnm) y de diversos tipos de vegetación (bosques tropicales caducifolios y subcaducifolios, bosques de pino, bosques de encino y bosques templados mixtos). En contraste, la distribución del lince se restringió a los bosques más fríos, localizados entre 2 000 y 2 800 msnm, una situación similar a la reportada en regiones cercanas como el estado de Oaxaca (Monroy & Briones- Salas, 2012).
Si bien hubo una alta coincidencia en los patrones de distribución de los felinos de afinidad tropical, existió también variación entre especies. Por ejemplo, mientras que alrededor del 40% de los registros del puma, ocelote y jaguarundi se presentaron en el bosque tropical caducifolio, solo el 30% de los registros de tigrillo, y alrededor del 20% de los de jaguar, se obtuvieron en este hábitat. Algo similar se observó en el caso del bosque de pino-encino, en donde existieron registros de todas las especies, siendo el ocelote la especie con el mayor porcentaje de registros con 57%, seguido del puma con 27%, el tigrillo con 12%, el jaguarundi con 2.5%, el jaguar con 1% y lince con 0.5%. Esto sugiere la existencia de factores que operan a escalas más finas que pueden influir de manera diferencial sobre las preferencias de los felinos por distintos tipos de hábitat. Por ejemplo, la importancia de los bosques templados de encino-pino de la región para los felinos de afinidad tropical, ocelote, tigrillo, jaguarundi y jaguar, puede deberse a que proporcionan recursos claves durante la época seca (p. ej., agua, alimento o refugio) en la que estos se vuelven muy escasos en las zonas más cálidas y secas. Esta posibilidad adquiere mayor apoyo cuando se considera que los registros de las cámaras-trampa obtenidos en esos bosques correspondieron, principalmente, a la época seca del año, y que existe evidencia que muestra que los manantiales son sitios donde se concentra la actividad de los felinos y sus presas en la región (Charre-Medellín, 2012).
En este sentido, es de destacar que el jaguarundi fue la especie cuyos registros se distribuyeron en el ámbito más amplio de variación climática a pesar de presentar un número reducido de registros. En contraste, el ocelote, a pesar de ser la especie con el mayor número de registros, presentó una concentración de los mismos en un intervalo de amplitud climática muy reducido. Por otra parte, algunos de los registros de jaguarundi, tigrillo y ocelote obtenidos en este estudio corresponden a las localidades más áridas y calientes en las que se ha registrado la presencia de poblaciones de esas especies de felinos en México. De esta manera los resultados de este estudio permiten tener una visión más amplia sobre la adaptabilidad de los felinos para habitar en un amplio ámbito de condiciones climáticas (Torres-Romero, 2009; Chávez-Tovar, 2010).
No sólo las condiciones climáticas determinan la abundancia y distribución de las especies. Para los felinos de mayor tamaño como el jaguar y puma, se ha documentado que prefieren hábitats con cobertura vegetal densa, poco fragmentada, distante de caminos, cercanos a cuerpos de agua y mínima perturbación humana en general (Thatcher, Van Manen, & Clark, 2006; Laundré, Loredo, Hernández, & Núñez, 2009; Charre-Medellín, 2012). En concordancia con esto, en este estudio se encontró que la distribución y la mayor cantidad de registros de puma y jaguar coincidieron con las áreas con una mayor extensión continua de bosques tropicales y templados, poca densidad humana y disponibilidad de presas (Charre-Medellín, 2012). Por otra parte, los registros obtenidos en sitios agropecuarios, correspondieron principalmente a casos en los que los habitantes locales reportaban felinos acechando tanto al ganado o a aves de corral. Un estudio más detallado sobre el efecto del impacto humano en las poblaciones de felinos, puede servir para entender con mayor detalle cómo la perturbación humana interactúa con el clima y la escasez de recursos claves, como el agua, que pueden influir en los patrones de distribución y abundancia de los felinos, en un sistema altamente heterogéneo como el presente en el occidente de México.
Por otra parte, conocer la distribución de las especies a nivel regional, es el primer paso para implementar acciones de conservación, que permitan la persistencia de las poblaciones locales de especies, principalmente, aquellas que por sus requerimientos son más vulnerables a la extinción local, caso es el de los carnívoros en general y felinos en particular (Kanagaraj, Wiegand, Mohamed, & Kramer-Schadt, 2013). En el área de estudio, urge contar con este tipo de información porque los bosques tropicales y bosques templados están actualmente afectados por elevadas tasas de deforestación anual de 1 y 1.8%, además de una tasa común de degradación del 20% en la extensión total de cubiertas de bosques y selvas (Bocco et al., 2001; Trejo & Dirzo, 2000; Mas-Caussel, Velásquez-Mon-tes, & Fernández-Vargas, 2005).
A esto se agrega la baja cobertura de áreas protegidas en las regiones costeras y la Sierra Madre del Sur, donde los resultados indican que se encuentra la mayor cantidad de registros de felinos. El presente estudio, sumado a posteriores iniciativas, que continúen documentando la presencia de felinos en zonas poco exploradas de la costa del Pacífico, son particularmente necesarias para delimitar nuevas áreas de conservación de felinos en: áreas del Bajo Balsas que colindan con el Estado de México y Guerrero en el Este, los bosques secos del Noroeste del Bajío, Este del Eje Neovolcánico y la Sierra Madre del Sur. Estas nuevas coberturas permitirían incrementar la conectividad entre poblaciones, para establecer una red con las pocas áreas naturales protegidas existentes del centro y occidente del país como Jalisco y el Estado de México (Monroy-Vilchis, Sánchez, Aguilera-Reyes, Suarez, & Urios, 2008; Aranda, Botello, & López-de Buen, 2012). De esta manera, la exploración de todas estas áreas, permitirá seguir incrementando el conocimiento sobre los factores que determinan los patrones de distribución de felinos a lo largo de gradientes ecológicos, permitiendo contar con mayores elementos para enfrentar el impacto humano actual y amenazas futuras como el cambio climático, el cual se anticipa tendrá un impacto significativo en la región al aumentar la temperatura promedio (3.6°C) y disminuir la precipitación anual promedio (-7.8%) en las próximas décadas (Sáenz-Romero et al., 2010; Sáenz-Romero et al., 2012)