











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versão On-line ISSN 0034-7744versão impressa ISSN 0034-7744
Rev. biol. trop vol.60 no.3 San José Set. 2012
Distribución espacial de Ocypode quadrata (Decapoda: Ocypodidae) en ocho playas de la zona norte oriental de Cuba
*Dirección para correspondencia
Abstract
Spatial distribution of Ocypode quadrata (Decapoda: Ocypodidae) in eight beaches of NorthEastern Cuba. Studies on the ecology of Ocypode quadrata have been mostly carried out in the Northern and Southern part of its distribution range. In despite that this species is common in Cuban beaches, there are no quantitative studies regarding its abundance and spatial distribution. The aim of this study was to report some aspects about the spatial variation of O. quadrata density in sandy beaches, with different levels of human influence, in the North coast of Eastern Cuba. For this, on May 2010, eight beaches with different levels of human influence were surveyed. On each beach, the number of crabs burrows were counted in 45 quadrats of 4m2 located in three different strata (P1, P2 and P3). According to burrow opening diameter, crabs were separated into young and adults forms. To determine the existence of statistical differences in the density of crab burrows among beaches and strata, a two-way ANOVA was developed with a Scheffé-procedure post hoc test. A total of 355 burrows were counted in 360 sample units. The composition by size classes was 237 burrows for young and 118 for adults. From the total of burrows, 74% were located in P1, 20% in P2 and 6% in P3. The higher concentration of burrows was found at Jiguaní beach (0.52±0.08 burrows/m2) while the lesser concentration was found at Estero Ciego beach (0.06±0.01 burrows/m2). Most of the beaches did not present significant differences in the burrows density (Scheffé, p>0.05), according to ANOVA results, in despite their different human influence level. Density of individuals was significantly higher in the upper intertidal (P1) areas (Scheffé, p<0.05) with predominance of young crabs. Total density diminished in P2 and P3 strata where a predominance of adult individuals was observed. The interaction term of beach and strata evidenced that the pattern of variation among strata was not the same for all beaches. The general pattern of adults and young specimen spatial distribution in the beaches was very similar to those reported in the literature. In the present study, the first conducted in Cuban beaches, we did not find evidence of direct relationship between the level of human impact and the density of ghost crab burrows.
Key words: Ocypode quadrata, sandy beach, zonation, human impact, Cuba.
Resumen
Uno de los organismos más comunes en las playas arenosas son los miembros del género Ocypode conocidos como cangrejos fantasmas, están ampliamente distribuidos en las regiones tropicales y subtropicales; además tienen un importante papel en la transferencia de energía en diferentes niveles tróficos. El objetivo del presente trabajo es dar a conocer aspectos relacionados con la variación espacial de la densidad de O. quadrata en playas con diferente nivel de influencia humana en la costa norte oriental de Cuba. En mayo de 2010, fueron contadas un total de 355 galerías en 360 unidades de muestreo. La composición por clases de tallas fue de 237 galerías de jóvenes y 118 de adultos. Del total de galerías contadas el 74% ocuparon el nivel P1, 20% el nivel P2 y 6% el nivel P3. La densidad varió entre 0.06-0.52galerías/m2. A pesar del nivel de influencia humana, la mayoría de las playas no mostró diferencias significativas en la densidad de galerías (Scheffé, p>0.05). La densidad de individuos en la zona superior de resaca del oleaje es mayor (Scheffé, p<0.05), predominando los jóvenes y en el resto de los estratos la densidad total va disminuyendo y predominan los individuos adultos. El patrón de variación entre los estratos no fue el mismo para todas las playas. En el presente estudio no se encontró evidencia directa entre el nivel de turismo y la densidad de galerías del cangrejo fantasma.
Palabras clave: Ocypode quadrata, playa arenosa, zonación, impacto humano, Cuba.
Las playas arenosas expuestas son los ambientes marinos que están más estructurados por factores físicos (McLachlan 1990). Una revisión reciente enfocada en la ecología de las playas arenosas, mostró un campo emergente de investigación que toma forma alrededor de un paradigma central y varias hipótesis bajo prueba (Defeo & McLachlan 2005, McLachlan & Brown 2006) donde se relaciona el modelo morfodinámico, propuesto por Short & Wright (1983), con diferentes aspectos de la fauna a distintos niveles y escalas.
Globalmente, el intenso crecimiento poblacional en las áreas costeras está conllevando a fuertes presiones sobre las playas a escalas y amplitudes sin precedentes (Brown & McLachlan 2002). Estos ecosistemas muestran una amplia plasticidad como respuesta a las variaciones en el clima, pero las modificaciones humanas de la zona costera limitan esta flexibilidad. Las limitaciones de nuestra comprensión científica de cómo las playas responden ecológicamente ante la gama de amenazas humanas están emergiendo rápidamente como impedimentos cruciales para la conservación de estos ecosistemas tan amenazados (Schlacher et al. 2007a).
Uno de los organismos más comunes en las playas arenosas son los miembros del género Ocypode conocidos como cangrejos fantasmas, que están ampliamente distribuidos en las regiones tropicales y subtropicales (Brown & McLachlan 1990). En el Atlántico occidental están representados por la especie Ocypode quadrata (Fabricius 1787), que se distribuye desde Rhode Island en Estados Unidos hasta Río Grande del Sur en Brasil (Melo 1996). Viven en galerías que construyen en la zona supralitoral desde la zona superior de resaca del oleaje hasta las dunas (Rathbun 1818).
Este organismo tiene un importante papel en la transferencia de energía en diferentes niveles tróficos pues, además de sus hábitos alimentarios como depredador y carroñero (Wolcott 1978), también puede extraer las microalgas que se encuentran en los sedimentos (Robertson & Pfeiffer 1982). En playas de Brasil se han documentado aspectos relacionados con la distribución espacial de O. quadrata (Turra et al. 2005, Neves & Bemvenuti 2006, Rosa & Borzone 2008, Araujo et al. 2008) y con la variación espacio-temporal de su abundancia, así como la influencia de los depredadores sobre la estructura poblacional de la especie (Branco et al. 2010). En México, se han realizado estudios sobre su abundancia y los patrones diarios de actividad (Valero-Pacheco et al. 2007).
O. quadrata, al igual que otras especies de su género, se ha usado como un indicador de los disturbios ocasionados por los humanos en las playas. Las menores densidades del cangrejo fantasma se han observado en zonas afectadas por el paso constante de las personas (Blankensteyn 2006, Neves & Bemvenuti 2006, Araujo et al. 2008), el tráfico de vehículos motorizados (Moss & McPhee 2006, Schlacher et al. 2007b), la construcción de obras civiles (Barros 2001), así como el relleno artificial de playas (Peterson et al. 2000).
La mayoría de los estudios sobre la ecología de O. quadrata se han llevado a cabo en los extremos de su ámbito de distribución a excepción de los trabajos realizados por Araujo et al. (2008) y por Valero-Pacheco et al. (2007) en playas tropicales. A pesar que en Cuba se reconoce a O. quadrata como un organismo común en las playas (Martínez-Iglesias 2007), aún no se ha realizado ningún estudio sobre la abundancia y distribución de esta especie. El objetivo del presente trabajo es dar a conocer aspectos relacionados con la variación espacial de la densidad de O. quadrata en playas con diferentes niveles de influencia humana en la costa norte oriental de Cuba.
Materiales y métodos
Área de estudio: La costa norte oriental de Cuba se caracteriza por tener una plataforma muy estrecha que alcanza los 100m de profundidad a una distancia menor de un kilómetro a partir de la línea de costa, excepto en las zonas donde existen bajos fondos y crestas coralinas que llegan a alcanzar una amplitud de dos kilómetros. Se distinguen dos tipos de costas, las acumulativas y las abrasivas, que están compuestas por diversos biotopos marinos. Entre las primeras se encuentran las de arenas que conforman las playas y las biogénicas cenagosas con esteros. Las costas arenosas en su mayoría son biogénicas y se ubican fundamentalmente en playas expuestas, las playas de arenas terrígenas son las menos frecuentes y se ubican hacia la desembocadura de algunos ríos o en el interior de algunas bahías. De forma general las playas expuestas no presentan gran extensión y se alternan con costas bajas de mangles o con costas rocosas bajas. En esta zona el régimen de marea predominante es semi-diurno con una amplitud media de 0.6m (Rodríguez & Córdova 2006). Se distinguen dos épocas climáticas: lluvias (mayo-octubre) y seca (noviembre-abril).
Sitios de muestreo: Para este estudio se seleccionaron ocho playas con diferentes niveles de influencia humana. Desde el punto de vista morfodinámico estas playas se clasifican como intermedias (Indice de Dean Ω=2.04-3.95) con una tendencia a ser reflectivas. Las playas se ordenaron de oeste a este en tres categorías según la intensidad de uso de las mismas: A playas de uso en temporada de verano y fines de semana y que presentan instalaciones recreativas; B playas de uso permanente, donde existen instalaciones hoteleras; C playas donde el uso por los bañistas es muy esporádico y que no presentan instalaciones de ningún tipo. Según el nivel de impacto, una clasificación ordinal cualitativa de estas categorías (Turra et al. 2005) tiene el siguiente orden: B>A>C.
En la categoría A se encuentran playa Blanca (21°05’03” N - 76°00’23” W) y Don Lino (21°05’18” N-75°59’45” W) compuestas por arenas biogénicas; en la primera playa, la parte superior está ocupada por árboles de la especie Coccoloba uvifera con escasa cobertura de plantas rastreras, mientras que la segunda playa está ocupada por la planta herbácea Ipomea prescaprae. En la categoría B, se encuentran Estero Ciego (21°06’47” N - 75°52’23” W), Atlántico (21°07’17” N - 75°50’22” W) y Las Brisas (21°07’33” N - 75°49’50” W), estas dos últimas ubicadas dentro del sector costero conocido como Guardalavaca; estas playas también tienen arenas biogénicas, presentan una vegetación compuesta fundamentalmente por árboles sin la presencia de una duna consolidada, limitando hacia tierra con un acantilado rocoso en el caso de las dos primeras y por un paseo peatonal en el caso de Las Brisas. En la categoría C se encuentran las playas Fundadora (20°33’33” W - 74°42’60” W), Jiguaní (20°32’15” N - 74°40’39” W) y Mapurisí (20°29’14” N - 74°36’34” W), donde la génesis de las arenas es terrígena, mixta y biogénica, respectivamente; en las dos primeras playas existe una duna baja bien consolidada compuesta por árboles y plantas herbáceas y rastreras, mientras que en Mapurisí la duna está ocupada por árboles de las especies Coccus nucifera y Coccoloba uvifera con algunos parches de la planta herbácea Sporobolus sp. Estas últimas tres playas se encuentran ubicadas dentro del Parque Nacional Alejandro de Humboldt. Durante el periodo de muestreo hubo condiciones climáticas muy buenas en todas las playas.
Muestreo: Se llevó a cabo un muestreo instantáneo en cada playa entre el 12-27 de mayo 2010, siempre temprano en la mañana desde las 6:00-9:00 horas. En cada playa se seleccionaron tres sectores (uno en cada extremo y uno en el centro). En cada sector se delimitaron tres estratos: P1- parte superior de la zona de resaca del oleaje, P2- berma y P3- límite de la vegetación de la duna o de las infraestructuras. En cada estrato se delimitó una parcela de 100m2 (50x2m) dentro de la cual se seleccionaron cinco réplicas de 4m2 (2x2m), considerada la unidad de muestreo, para un total de 45 unidades de muestreo por playa. En cada unidad de muestreo se contó el número de galerías, este censo indirecto del número de individuos se ha empleado en otros estudios (Wolcott 1978, Fisher & Tevesz 1979, Warren 1990, Alberto & Fontoura 1999, Barros
2001, Turra et al. 2005, Moss & McPhee 2006, Neves & Bemvenuti 2006). Se consideraron como galerías ocupadas aquellas en las que se evidenció la existencia de actividad de excavación debido a la presencia de montículos de arena húmeda.
El diámetro de la abertura de cada galería se midió con un calibre digital (0.01mm) también como una medida indirecta de la talla (Wolcott 1978, Fisher & Tevesz 1979, Alberto
& Fontoura 1999, Turra et al. 2005). El tamaño de la abertura de la galería se separó en dos clases de tallas: jóvenes (menor de 25mm) y adultos (mayor de 25mm). Alberto & Fontoura (1999) propusieron que los diámetros de las galerías a partir de 23mm podían considerarse como individuos que ya habían alcanzado su madurez sexual.
La densidad (número medio de galerías/ m2±error estándar) se calculó para cada uno de los estratos en cada playa. Para comparar el número de galerías entre playas y entre los diferentes estratos, se usó un Análisis de Varianza (ANOVA) de dos factores. Para cumplir con los requisitos de la distribución normal todos los datos fueron transformados a la raíz cuadrada de (x+½). Los resultados del ANOVA fueron seguidos por una prueba de Scheffé para realizar comparaciones a posteriori. Las diferencias estadísticas fueron consideradas a un nivel de significación de 95% (α=0.05) (Zar 1999).
Resultados
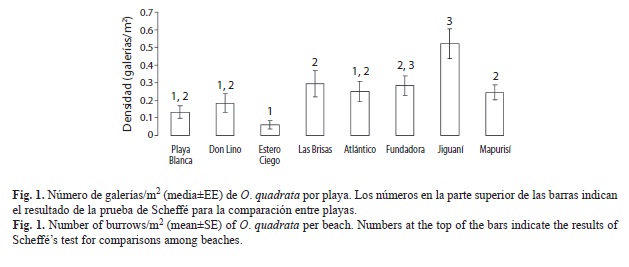
Fueron contadas un total de 355 galerías en 360 unidades de muestreo. La composición por clases de tallas fue de 237 galerías de jóvenes y 118 de adultos. La mayor densidad de galerías fue encontrada en playa Jiguaní (0.52±0.08galerías/m2), mientras que la menor densidad se registró en Estero Ciego (0.06±0.01galerías/ m2). En el resto de las playas la densidad promedio varió entre 0.13- 0.29galerías/m2 (Fig. 1). Los resultados del ANOVA muestran que en general existen diferencias significativas en la densidad de galerías entre las playas (F=11.415, g.l.=7, p=0.0000), sin embargo la prueba a posteriori de Scheffé arrojó que algunas playas no difieren de otras con independencia del nivel de impacto que presenten. En la figura 1 se evidencian las diferencias entre Estero Ciego (categoría B) y Jiguaní (categoría C) formando dos grupos independientes; sin embargo, existe un tercer grupo formado por playa Blanca (A), Don Lino (A), Las Brisas (B), Atlántico (B), Fundadora (C) y Mapurisí (C), cuya densidad no difiere significativamente entre ellas a pesar de tener diferentes niveles de influencia humana. Este último grupo a su vez puede presentar playas que no difieren en densidad con respecto a alguno de los dos primeros grupos, así playa Blanca (A), Don Lino (A) y Atlántico (B) no difieren de Estero Ciego (B), y Fundadora (C) no difiere de Jiguaní (C).
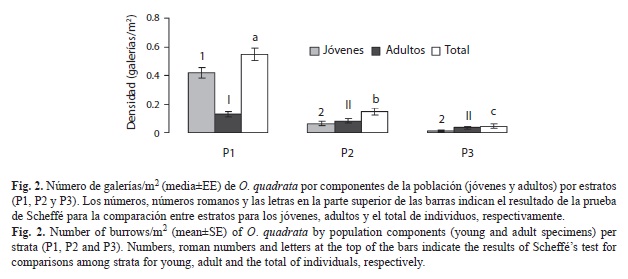
Del total de galerías contadas el 74% ocuparon el estrato P1, 20% el estrato P2 y 6% el estrato P3, existiendo diferencias significativas en la densidad de galerías entre los estratos (F=18.287, g.l.=2, p=0.0000), este resultado fue corroborado por la prueba de Scheffé (Fig. 2). Al sumar todos los datos por playas para determinar si existe zonación entre los diferentes componentes de la población, se observó que desde el estrato P1-P3 va disminuyendo la densidad de galerías pertenecientes a individuos jóvenes y adultos. En todos los estratos se encontraron galerías de individuos jóvenes y adultos pero la proporción cambió en los dos estratos superiores. Mientras que en P1 existe mayor densidad de jóvenes, en P2 y P3 es mayor la densidad de adultos. Para cada componente de la población se realizó un ANOVA unifactorial (α=0.05) (Zar 1999) mostrando que el estrato P1 difiere significativamente de P2 y P3 en cuanto a la densidad de galerías tanto para individuos jóvenes (Scheffé, p<0.05) como para individuos adultos (Scheffé, p<0.05) pero los dos últimos estratos no difieren uno de otro (Fig. 2).
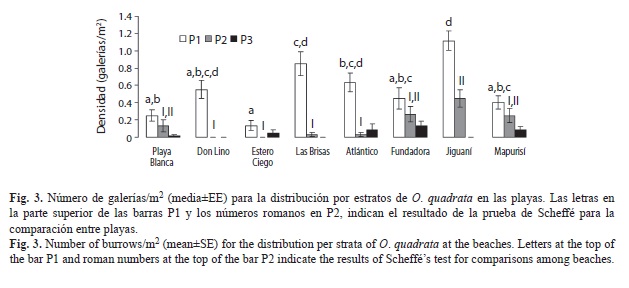
La interacción entre los factores playa y estrato, evidenció que el patrón de variación entre estratos no fue el mismo para todas las playas (F=7.533, g.l.=14, p=0.0000). En todas las playas la mayor densidad media de galerías se encontró en la parte superior de la zona de resaca del oleaje, alcanzando los máximos valores en Jiguaní (C) (1.12±0.06galerías/m2) y en Las Brisas (B) (0.85±0.14 galerías/m2); los valores mínimos se encontraron en Estero Ciego (B) (0.13±0.06galerías/m2) y en Don Lino (A) (0.25±0.06galerías/m2). La densidad en la berma y en el límite superior de la playa siguió un patrón diferente: en Don Lino (A) no se encontraron galerías en los estratos P2 y P3, en Estero Ciego (B) no se encontraron en P2 y en Las Brisas (B) y Jiguaní (C) no aparecieron galerías en el estrato P3 (Fig. 3).
Teniendo en cuenta la complejidad del análisis a partir de la densidad media calculada para los tres estratos, que las características del estrato P3 varía entre las playas (ver descripción de los sitios de muestreo) y que la presión de uso de estas playas se realiza en los dos primeros estratos, se decidió realizar un ANOVA de un factor (α=0.05) (Zar 1999) comparando las playas de forma independiente para P1 y P2. En el estrato P1 existen diferencias entre las playas (F=11.004, g.l.=7, p=0.0000), separándose las playas Estero Ciego (B) y Jiguaní (C) en dos grupos completamente diferentes y otros dos grupos conformados por el resto de las playas con independencia del nivel de influencia humana presente (Fig. 3). En el estrato P2 también existen diferencias entre las playas (F=8.012, g.l.=7, p=0.0000), existiendo dos grupos, el primero formado por Estero Ciego (B), Don Lino (A), Las Brisas (B) y Atlántico (B) y el segundo por Playa Blanca (A), Fundadora (C), Mapurisí (C) y Jiguaní (C). Las tres primeras playas de este segundo grupo no difieren significativamente con las del primero, por lo que nuevamente se observa independencia en la densidad con respecto al nivel de influencia humana (Fig. 3).
Discusión
A pesar de abarcar solo una época climática, este es el primer reporte que cuantifica la abundancia y describe la distribución espacial de O. quadrata en playas de Cuba. La densidad de galerías de O. quadrata en las playas de la costa norte oriental fue, de forma general, menor que la encontrada por Turra et al. (2005), Neves & Bemvenuti (2006), Rosa & Borzone (2008) en las costas de Río Grande del Sur (Brasil) y por Valero-Pacheco et al. (2007) en una playa de Veracruz (México). En una playa del sur de Brasil fueron reportados valores de densidades entre 0.06-0.7galerías/ m2 (Alberto & Fontoura 1999) mientras que en playas al norte se registró una densidad entre 0.08-0.33galerías/m2 (Araujo et al. 2008), valores similares a los obtenidos en el presente estudio. En las costas de Estados Unidos se encontraron valores de densidad entre
0.005-0.014galerías/m2 (Wolcott & Wolcott 1984), mientras Hobbs et al. (2008) encontraron valores entre 0.02-0.13galerías/m2. Defeo & Cardoso (2004) en un análisis latitudinal del rango de distribución de Emerita brasiliensis, atribuyeron que las variaciones en la abundancia se deben a variaciones locales en parámetros físicos (morfodinámica y grado de exposición) y biológicos (competencia) en lugar de los efectos biogeográficos.
El patrón general de distribución de adultos y jóvenes en las playas es muy similar a los informados para la misma especie en diferentes países. La mayor densidad de galerías en la zona superior de resaca del oleaje coincide con lo reportado por Leber (1982), Hill & Hunter (1973), Turra et al. (2005) y Branco et al. (2010) para ambos extremos del ámbito de distribución de la especie y para playas tropicales en México (Valero-Pacheco et al. 2007) y en Brasil (Araujo et al. 2008). La distribución diferencial en un gradiente perpendicular de la playa parece estar relacionada con el desarrollo ontogenético de la especie (Turra et al. 2005), hecho que ha tratado de ser explicado por aspectos como una menor resistencia de los jóvenes a la desecación y menor capacidad de construir galerías profundas en zonas superiores de la playa (Fisher & Tevesz 1979). La segregación por tallas ha sido documentada para anfípodos, isópodos, cangrejos hipoideos y almejas (Defeo et al. 1986, Jaramillo et al. 1994, McLachlan & Jaramillo 1995, Cardoso & Veloso 2003, Ocaña et al. 2010) y sugiere una capacidad diferencial de cada componente de la población para seleccionar un microhabitat deseado, así como un mecanismo evolutivo para evitar competir por espacio y alimento. Por otra parte se conoce que O. quadrata es una especie territorialista (Fisher & Tevesz 1979) y tal vez a una densidad determinada en la zona superior de resaca del oleaje, los individuos comienzan a ocupar la berma y la duna.
La segregación por estratos de la playa también pueden influir en aspectos relacionados con las características específicas de cada playa tales como: la competencia intraespecífica o la mayor disponibilidad de recursos. Jiguaní, que es una playa con la presencia de todos sus elementos morfológicos y es muy prístina, presenta una duna con una densa vegetación compuesta por la planta herbácea Ipomea prescaprae que pudiera ser responsable de la ausencia de galerías por ocupar totalmente el sustrato. En Mapurisí, la duna está ocupada por algunos parches de la planta Sporobolus sp., dejando espacios del sustrato libre en los que los cangrejos pueden cavar sus galerías. Otras playas, como Don Lino y Las Brisas, no presentaron galerías en el estrato superior tal vez debido a que en esta parte en ambas playas se evidencia mayor compactación de la arena.
Muchas de las especies de la macrofauna de playas arenosas se manifiestan como buenos indicadores para medir el impacto sobre estos ecosistemas (Defeo et al. 1986, Defeo & de Álava 1995, Lercari & Defeo 1999). Varios estudios han tratado de presentar el nivel de influencia humana como un factor que puede determinar, o al menos influir en, los cambios en la distribución del género Ocypode a diferentes escalas espaciales. Barros (2001), trabajando con O. cordimana, y Neves & Bemvenuti (2006) y Araujo et al. (2008), con O. quadrata, encontraron que la densidad promedio de galerías disminuyó desde zonas sin impacto humano a zonas con alta ocupación del litoral. Turra et al. (2005) también sugirieron este tipo de relación aunque sus resultados no fueron significativos desde el punto de vista estadístico en cuanto a plantear un modelo lineal entre densidad de galerías y nivel de turistas en playas micromareales de Brasil.
En el presente estudio no se encontró evidencia de relación directa del nivel impacto con la densidad de galerías del cangrejo fantasma. Si bien una de las playas más ocupadas presentó la menor densidad y otra muy prístina presentó los mayores valores de densidad promedio de galerías por unidad de muestreo, el resto de las playas no presentó, de forma general, diferencias entre ellas, independientemente del nivel de impacto en cada una. En el análisis entre los dos primeros estratos de la playa para descartar la influencia de la presencia y estado de las dunas, los resultados fueron diferentes. En el primer estrato (P1) existieron playas con diferentes niveles de influencia que no presentaron diferencias entre sí, sin embargo, en el segundo estrato (P2) las playas ubicadas en el Parque Nacional mostraron mayores valores de densidad de galerías que las restantes. El estrato P2 podría ser la zona que más influencia de actividades tiene en toda la playa. Es común encontrar instalaciones ligeras ubicadas en la berma, es un lugar de la playa ideal para realizar actividades recreativas tal como juegos, además es un lugar con mayor frecuencia de acciones de limpieza.
Gheskiere et al. (2005), demostraron que la zona supralitoral de las playas arenosas usadas por los turistas al norte y sur de Europa, tienen menor concentración de materia orgánica y menor abundancia y diversidad de invertebrados en comparación con las playas cercanas que no tienen influencia de turistas. Este hecho lo atribuyen a la combinación de acciones de limpieza de playas, al pisoteo y otros disturbios directos ocasionados por los usuarios. Con base en otras actividades recreativas en playas como el uso de vehículos, se encontraron menores valores de densidad de galerías del cangrejo fantasma (Moss & McPhee 2006) y mayores valores de mortalidad (Schlacher et al. 2007b) en playas con mayor tráfico.
Para los macroinvertebrados se ha encontrado que la abundancia sigue un patrón relacionado con la morfodinámica (Defeo & McLachlan 2005, Defeo & McLachlan 2011). En este sentido se ha encontrado que, para especies supralitorales como los anfípodos (Defeo & Gómez 2005) y O. quadrata (Defeo & McLachlan 2011), la abundancia disminuye de playas reflectivas a disipativas. Las playas estudiadas en la zona norte oriental de Cuba se clasifican en un estado intermedio por lo tanto, con base en las predicciones del modelo morfodinámico (Defeo & McLachlan 2005, Defeo & McLachlan 2011), sería de esperar que la abundancia de galerías no presentara diferencias entre ellas. Las relaciones interespecíficas también pueden llegar a influir sobre la estructura de una población de O. quadrata. Branco et al. (2010) encontraron que la presión ejercida por depredadores puede ser significativa en la disminución de la abundancia.
Algunos de los trabajos realizados para tratar de relacionar el nivel de influencia humana con parámetros poblacionales como la abundancia y la estructura de tallas del cangrejo fantasma, han tenido la limitante de basarse en muestreos instantáneos (Barros 2001, Turra et al. 2005, Neves & Bemvenuti 2006). Otro elemento a tener en cuenta es el procedimiento en el análisis de los datos. Barros (2001) no encontró diferencias entre tres playas que se ubicaban en una zona urbanizada; de forma independiente comparó seis playas en una zona no urbanizada y encontró diferencias significativas entre las mismas. Este mismo autor comparó los datos acumulados para las dos categorías mediante un ANOVA y encontró diferencias significativas de la densidad del cangrejo fantasma entre playas urbanizadas y no urbanizadas.
Siguiendo este mismo análisis en las playas objeto del presente estudio, se observó que la densidad de galerías mostró diferencias (Scheffé, p<0.05) entre Las Brisas y Estero Ciego (categoría B) y entre Jiguaní y Mapurisí (categoría C), lo que puede interpretarse como que el nivel de influencia humana no es el factor determinante en las diferencias de la densidad de galerías entre las playas sino que debe haber otros factores actuando en estrecha relación. Es recomendable diseñar estudios que logren introducir todas las variables dentro de un mismo modelo para dilucidar la contribución relativa de cada factor en modular la dinámica poblacional de esta especie. Este tipo de correlación puede llevarse a cabo bajo diseños de muestreo bien concebidos, trabajando a diferentes escalas y con réplicas y controles bien planificados (Turra et al. 2005).
Agradecimientos
Agradecemos la colaboración de Geovanis Rodríguez Cobas y al personal del Parque Nacional Alejandro de Humboldt por su ayuda durante el trabajo de campo. A Carlos Peña por su apoyo logístico. A Paulo Lana por motivarnos a la realización de este trabajo y por facilitarnos toda la literatura.
Referencias
Alberto, R.M.F. & N.F. Fontoura. 1999. Distribuição e estrutura etária de Ocypode quadrata (Fabricius, 1787) (Crustacea, Decapoda, Ocypodidae) em praia arenosa do litoral sul do Brasil. Rev. Brasil. Biol. style="font-family: verdana;">59: 95-108. [ Links ]
Araujo, C.C.V., D.M. Rosal & J.M. Fernandes. 2008. Densidade e distribuição espacial do caranguejo Ocypode quadrata (Fabricius, 1787) (Crustacea, Ocypodidae) em três praias arenosas do Espírito Santo, Brasil. Biotemas 21: 73-80. [ Links ]
Barros, F. 2001. Ghost crabs as a tool for rapid assessment of human impacts on exposed sandy beaches. Biol. Conservat. 97: 399-404. [ Links ]
Blankensteyn, A. 2006. O uso do caranguejo maria-farinha Ocypode quadrata (Fabricius) (Crustacea, Ocypodidae) como indicador de impactos antropogênicos em praias arenosas da Ilha de Santa Catarina. Rev. Bras. Zool. 23: 870-876. [ Links ]
Branco, J.O., J.C. Hillesheim, H.A.A. Fracasso, M.L. Christoffersen & C.L. Evangelista. 2010. Bioecology of the ghost crab Ocypode quadrata (Fabricius, 1787) (Crustacea: Brachyura) compared with other intertidal crabs in the southwestern Atlantic. J. Shellf. Res. 29: 503-512. [ Links ]
Brown, A.C. & A. McLachlan. 1990. Ecology of Sandy Shores. Elsevier, Amsterdam, Holanda. [ Links ]
Brown, A.C. & A. McLachlan. 2002. Sandy shore ecosystems and the threats facing them: some predictions for the year 2025. Environ. Conservat. 29: 62-77. [ Links ]
Cardoso, R.S. & V.G. Veloso. 2003. Population dynamics and secondary production of the wedge clam Donax hanleyanus (Bivalvia: Donacidae) on a high-energy, subtropical beach of Brazil. Mar. Biol. 142: 153-162. [ Links ]
Defeo, O. & A. de Álava. 1995. Effects of human activities on long-term trends in sandy beach populations: the wedge clam Donax hanleyanus in Uruguay. Mar. Ecol. Prog. Ser. 123: 73-82. [ Links ]
Defeo, O. & A. McLachlan. 2005. Patterns, processes and regulatory mechanisms in sandy beach macrofauna: a multi-scale analysis. Mar. Ecol. Prog. Ser. 295: 1-20. [ Links ]
Defeo, O. & A. McLachlan. 2011. Coupling between macrofauna community structure and beach type: a deconstructive meta-analysis. Mar. Ecol. Prog. Ser. 433: 29-41. [ Links ]
Defeo, O. & J. Gómez. 2005. Morphodynamics and habitat safety in sandy beaches: life-history adaptations in a supralittoral amphipod. Mar. Ecol. Prog. Ser. 293: 143-153. [ Links ]
Defeo, O. & R. Cardoso. 2004. Latitudinal patterns in abundance and life-history traits of the mole crab Emerita brasiliensis on South American sandy beaches. Diversity and Distributions 10: 89-98. [ Links ]
Defeo, O., C. Layerle & A. Masello. 1986. Spatial and temporal structure of the yellow clam Mesodesma mactroides (Deshayes, 1854) in Uruguay. Medio Ambiente (Chile) 8: 48-57. [ Links ]
Fisher, J.B. & M.J.S. Tevesz. 1979. Within-habitat patterns of Ocypode quadrata (Fabricius) (Decapoda Brachyura). Crustaceana 5: 31-36. [ Links ]
Gheskiere, T., M. Vincx, J.M. Weslawski, F. Scapini & S. Degraer. 2005. Meiofauna descriptor of tourisminduced changes at sandy beaches. Mar. Environ. Res. 60: 245-265. [ Links ]
Hill, G.W. & R.E. Hunter. 1973. Burrows of the ghost crab Ocypode quadrata (Fabricius) on the Barrier Islands, southcentral Texas coast. J. Sediment. Petrol. 43: 24-30. [ Links ]
Hobbs, C.H.III., C.B. Landry & J.E.III. Perry. 2008. Assessing anthropogenic and natural impacts on ghost crabs (Ocypode quadrata) at Cape Hatteras National Seashore, North Carolina. J. Coast. Res. 24: 1450-1458. [ Links ]
Jaramillo, E., M. Pino, L. Filún & M. González. 1994. Longshore distribution of Mesodesma donacium (Bivalvia: Mesodesmatidae) on a sandy beach of the south of Chile. Veliger 37: 192-200. [ Links ]
Leber, K. 1982. Spatial pattern of Ocypode quadrata: a re-evaluation (Decapoda, Brachyura). Crustaceana 41: 110-112. [ Links ]
Lercari, D. & O. Defeo. 1999. Effects of freshwater discharge in sandy beach populations: the mole crab Emerita brasiliensis in Uruguay. Estuar. Coast. Shelf. Sci. 49: 457-468. [ Links ]
Martínez-Iglesias, J.C. 2007. Artrópodos Filo Arthropoda, Crustáceos Subfilo Crustacea: cangrejos, camarones, langostas y anomuros - Clase Malacostrata, Orden Decapoda, p. 73-77. In R. Claro (ed.). La Biodiversidad marina de Cuba. (CD-ROM). Instituto de Oceanología, Ministerio de Ciencia, Tecnología y Medio Ambiente, La Habana, Cuba. [ Links ]
McLachlan, A. 1990. Dissipative beaches and macrofauna communities on exposed intertidal sands. J. Coast. Res. 6: 57-71. [ Links ]
McLachlan, A. & E. Jaramillo. 1995. Zonation on sandy beaches. Oceanogr. Mar. Biol. Annu. Rev. 33: 305-335. [ Links ]
McLachlan, A. & A.C. Brown. 2006. The ecology of sandy shores. Academic, Massachusetts, EEUU. [ Links ]
Melo, G.A.S. 1996. Manual de identificação dos Brachyura (caranguejos e siris) do litoral brasileiro. Plêiade, São Paulo, Brasil. [ Links ]
Moss, D. & D.P. McPhee. 2006. The impacts of recreational four-wheel driving on the abundance of the ghost crab (Ocypode cordimanus) on a subtropical sandy beach in SE Queensland. Coast. Manag. 34: 133-140. [ Links ]
Neves, F.M. & C.E. Bemvenuti. 2006. The ghost crab Ocypode quadrata (Fabricius, 1787) as a potential indicator of anthropic impact along the Rio Grande do Sul coast, Brazil. Biol. Conserv. 133: 431-435. [ Links ]
Ocaña, F.A., A. Fernández, A. Silva, P.A. González & Y. García. 2010. Estructura poblacional de Donax striatus (Bivalvia, Donacidae) en playa Las Balsas, Gibara, Cuba. Rev. Mar. Cost. 2: 27-38. [ Links ]
Peterson, C.H., D.H.M. Hickerson & G.G. Johnson. 2000. Short term consequences of nourishment and bulldozing on the dominant large invertebrates of a sandy beach. J. Coast. Res. 16: 368-378. [ Links ]
Rathbun, M.J. 1818. The grapsoid crabs of America. Bull. US. Nat. Mus. Washington 97: 1-461. [ Links ]
Robertson, J.R. & W.J. Pfeiffer. 1982. Deposit-feeding by the ghost crab Ocypode quadrata (Fabricius). J. Exp. Mar. Biol. Ecol. 56: 165-177. [ Links ]
Rodríguez, R.A. & E.A. Córdova. 2006. La erosión en las playas de la región nororiental de Cuba. Revista Geográfica 139: 9-26. [ Links ]
Rosa, L.C. & C.A. Borzone. 2008. Spatial distribution of Ocypode quadrata (Crustacea: Ocypodidae) along estuarine environments in the Paranaguá Bay Complex, Southern Brazil. Rev. Bras. Zool. 25: 383-388. [ Links ]
Schlacher, T.A., J. Dugan, D.S. Schoeman, M. Lastra, A. Jones, F. Scapini, A. McLachlan & O. Defeo. 2007a. Sandy beaches at the brink. Diversity and Distributions 13: 556-560. [ Links ]
Schlacher, T.A., L. Thompson & S. Price. 2007b. Vehicles versus conservation of invertebrates on sandy beaches: mortalities inflicted by off-road vehicles on ghost crabs. Mar. Ecol. 28: 354-367. [ Links ]
Short, A.D. & L.D. Wright. 1983. Physical variability of sandy beaches, p. 133-144. In A. McLachlan & T. Erasmus (eds.). Sandy beaches as ecosystem. W. Junk, The Hague, Holland. [ Links ]
Turra, A., M.A.O. Gonçalves & M.R. Denadai. 2005. Spatial distribution of the ghost crab Ocypode quadrata in low-energy tide-dominated sandy beaches. J. Nat. Hist. 39: 2163-2177. [ Links ]
Valero-Pacheco, E., F. Álvarez, L.G. Abarca-Arenas & M. Escobar. 2007. Population density and activity pattern of the ghost crab, Ocypode quadrata, in Veracruz, Mexico. Crustaceana 80: 313-325. [ Links ]
Warren, J.H. 1990. The use of open burrows to estimate abundances of intertidal estuarine crabs. Aust. J. Ecol. 15: 277-280. [ Links ]
Wolcott, T.G. 1978. Ecological role of ghost crabs, Ocypode quadrata (Fabricius) on an ocean beach: scavengers or predators? J. Exp. Mar. Biol. Ecol. 31: 67-82. [ Links ]
Wolcott, T.G. & D.L. Wolcott. 1984. Impact of off-road vehicles on macroinvertebrates of a mid-Atlantic beach. Biol. Conserv. 29: 217-240. [ Links ]
Zar, J.H. 1999. Biostatistical Analysis. Prentice-Hall, Nueva Jersey, EEUU. [ Links ]
*Correspondencia:
Frank A. Ocaña: Centro de Investigaciones y Servicios Ambientales y Tecnológicos, Calle 18 s/n esq. a Maceo, Rpto. “El Llano”, Holguín, Cuba. franko@cisat.cu
Antonio Vega: Centro de Investigaciones y Servicios Ambientales y Tecnológicos, Calle 18 s/n esq. a Maceo, Rpto. “El Llano”, Holguín, Cuba. vega@cisat.cu
Elier A. Córdova: Centro de Investigaciones y Servicios Ambientales y Tecnológicos, Calle 18 s/n esq. a Maceo, Rpto. “El Llano”, Holguín, Cuba. elier@cisat.cu
1. Centro de Investigaciones y Servicios Ambientales y Tecnológicos, Calle 18 s/n esq. a Maceo, Rpto. “El Llano”, Holguín, Cuba; franko@cisat.cu, vega@cisat.cu, elier@cisat.cu
Recibido 24-VIII-2011. Corregido 10-III-2012. Aceptado 12-IV-2012.


{kind=link}
{kind=link}
{kind=link}