Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
The taxonomic classification of Masdevallia Ruiz & Pav. has been and still is com- plicated. Before molecular research was available to taxonomists, the classification was based on morpho- logical features. But once the “secrets” of the DNA molecule were revealed, multiple options how to split large orchid genera into smaller entities became avail- able. But regardless how “correct” this remarkable sci- entific development is, it still leaves plenty of room for subjective preferences in taxonomic treatments. In 2006, the genus Masdevallia had grown to include more than 500 species (Luer 2006), classified into numerous subdivisions (Luer 2000a,b, 2001, 2002, 2003). This vast number of species, in combination with newly available molecular evidence (Pridgeon & Chase 2001), encouraged Luer to split the genus into 16 new genera, in addition to the remaining Masdeval- lia (Luer 2006). However, the taxonomic advantages of this division of Masdevallia (Luer 2006) are ques- tionable and not recognized by the present and other authors, mainly due to the difficulties in separating the new genera from each other in consistent ways, and difficulties in identifying to which genus many species belong. Therefore, we favor the previous and more conservative taxonomic treatment of the genus as circumscribed by Luer (2000a,b, 2001, 2002, 2003), for scientific, user-friendly and practical reasons.
The genus Masdevallia is currently represented in Peru by approximately 198 recognized species, excluding the one described here (POWO 2023). The origin of the cultivated type specimen of this latter spe- cies is from a collecting trip in 2010, to the rich cloud forest east of Monopampa, Huanuco (Fig. 1). Several plants were collected in this area on that occasion and were introduced into cultivation and propagation by Peruflora, an officially authorized plant nursery estab- lished for the conservation and commercial utilization of native Peruvian plants, including orchids. When the cultivated Masdevallia plant flowered, it was com- pared to flowering plants recently observed and pho- tographed by the authors along the new road between Monopampa and Pozuzo (Fig. 2-4). The conclusion is that they are the same and represent a new species to science, which is described here.

Taxonomic treatment
Masdevallia emieliana Dalström, Deburghgr. & Ruíz Pérez, sp. nov. (Fig. 2, 3, 4, 5).
TYPE: Peru. Region of Huanuco: Province of Pachitea, District of Chaglia, Location: Millpo, along the ancient trail between Monopampa and Pozuzo, exact GPS co- ordinates for the original collection uncertain, flowered in cultivation 16 Nov, 2022 by Peruflora, S. Dalström 3942 (holotype: USM, accession N° 331222).
Diagnosis: Masdevallia emieliana appears to belong to the subgenus Pygmaeia, section Amaluzae, and seems to be most morphologically similar to M. fenestralis Dalström & Ruíz Pérez, and M. fenestrellata Dalström & Ruíz Pérez (Dalström & Ruíz Pérez 2015) which are sympatric. The new species differs from both by lacking the basal sepaline translucent fenestrations (“window- like” structures) and by the presence of external sepaline ridges. Although the basic general floral features of this new species are similar to many other Masdevallia species in other subgenera, the vegetative habit and the production of a second flower on the threadlike inflores- cence, however, are very similar to the above-mentioned sympatric species, which suggests a close relationship.
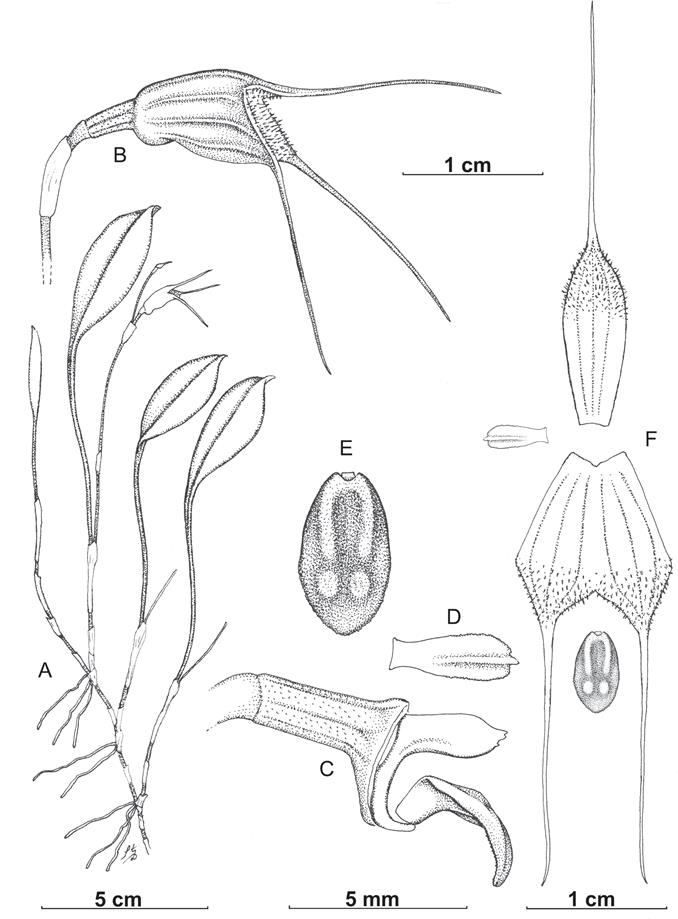
Epiphytic herb. Plant medium sized for the subge- nus, with a creeping rhizome with distant ramicauls ca. 2 cm apart, partially enclosed by one 5-6 mm long sheath. Ramicauls erect and rather thin, ca. 1.2-1.5 mm in di- ameter, ca. 30-35 mm long, enclosed basally by one to three 7-22 mm long tubular sheaths. Leaf erect to arch- ing, petiolate, slender, longitudinally furrowed, ca 1.5 mm in diameter and ca. 60 mm long, blade basally con- duplicate and cuneate, elliptic, with a broad and slightly folded apicule, 35-45 × 14-17 mm. Inflorescence erect, very thin and thread-like, ca. 1 mm in diameter, with a second successive flower on larger plants (like for the type), with an up to ca. 7 cm long peduncle; peduncular bract 1, tubular, ca. 6-7 mm long; floral bract appressed, tubular, 6-7 mm long; pedicel excluding the ovary ca. 8 mm long; ovary shallowly sulcate, smooth, with scattered tiny “fungal pits” (Dalström & Ruíz Pérez 2016), 2.4-2.5 mm long. Flower cupulate, forming a ca. 10 mm long sepaline cup; dorsal sepal whitish with purple irregular stripes along raised fleshy ridges, which follow the veins, internally similarly colored, but without furrows along the veins, apically covered with fine white hairs, ca. 13 × 5 mm, connate to the lateral sepals for 7-8 mm, then obtuse and terminated in a whitish, basally purple spot- ted ca. 18 mm long tail; lateral sepals similar in texture, hairiness and coloration, connate to the dorsal sepal, then obliquely acute, fused to each other for ca. 9-10 mm, and ca. 12 × 10 mm when measured combined, with apical, slender, whitish ca. 18-19 mm long tails; petals whitish, cartilaginous, indistinctly unguiculate with a distinct, lon- gitudinal, central, fleshy ridge emerging from the middle of the petal and ending in a fleshy protruding apicule, ca. 3.5 × 0.9-1.0 mm; lip whitish with purple center furrow and purplish hue apically, attached to the column foot by a short, strap-like “hinge”, lip lamina basally u-shaped then angled into an elliptic, flat, fleshy lamina, with two parallel longitudinal ridges, emerging from the base and ending near the middle of the lamina, then with a pair of low, fleshy knobs between the end of the ridges and the obtuse apex of the lamina, ca. 4.5 × 2.5 mm; column whitish, straight, ca. 2.7 mm long, apically indistinctly serrate, with an equally long, curved and apically hook- shaped foot; antercap and pollinia not seen.
Other specimens examined: Peru. Huanuco: Millpo, in wet cloud forest along the road between Monopampa and Pozuzo, S 09°48.123’; W 75°42.589’, alt. 2622 m, 16 Nov. 2022. Digital images only; Fig. 1-3 (Dalström archives). A few flowering plants were observed and photographed growing epiphytically on mossy branch- es. No other populations are known.
Distribution: Masdevallia emieliana is only known in the wild from the wet forests east of Monopampa, Huanuco, Peru.
Eponymy: This species is named in honor of Emiel Coppens of Nieuwerkerken near Aalst, Belgium, for being a great source of inspiration to the second author.

Photo by Stig Dalström.
Figure 2 Author Deburghgraeve inspects a blooming plant of what became Masdevallia emieliana.